Forskare har räknat antalet neuroner i den mänskliga hjärnan. Elektriska manifestationer av aktiviteten i hjärnbarken
Hemisfärernas cortex stor hjärnaär den högsta och mest komplexa organiserade nervcentrum skärmtyp, vars aktivitet säkerställer regleringen av olika funktioner i kroppen och komplexa former av beteende.
Barken bildas av ett lager av grå substans 3-5 mm tjockt. Den grå substansen innehåller nervceller (mer än 10 miljarder neuroner), nervfibrer och neurogliaceller. Dess olika delar, som skiljer sig från varandra i vissa särdrag av cellernas placering och struktur, placeringen av fibrer och funktionell betydelse, kallas fält, som först beskrevs av den tyske läkaren och vetenskapsmannen K. Brodmann.
Cytoarkitektonik
Bland de multipolära neuronerna i cortex urskiljs pyramidala, stellate, fusiforma, arachnid, horisontella, "kandelabra" celler, celler med en dubbel bukett av dendriter och några andra typer av neuroner.
Pyramidala neuroner utgör den huvudsakliga och mest specifika formen för hjärnbarken. De har en långsträckt konformad kropp, vars spets vetter mot ytan av cortex. Dendriter sträcker sig från apex och laterala ytor av kroppen. Axoner härstammar från basen av pyramidcellerna.
Pyramidceller av olika lager av cortex skiljer sig i storlek och har olika funktionellt värde. Små celler är interkalära neuroner. Axoner av stora pyramider deltar i bildningen motor pyramidformiga vägar.
Nervcellerna i cortex är belägna i oskarpt avgränsade lager, som betecknas med romerska siffror och numrerade från utsidan till insidan. Varje lager kännetecknas av dominansen av någon typ av cell. Det finns sex huvudlager i hjärnbarken:
- I - molekylär;
- II - extern granulär;
- III - pyramidal;
- IV - inre granulär;
- V - ganglion;
- VI - lager av polymorfa celler.
jag- Molekyl barkskiktet innehåller Ett stort antal små associativa horisontella Cajal-celler. Deras axoner löper parallellt med hjärnans yta som en del av det tangentiella plexus av nervfibrer i det molekylära lagret. Men huvuddelen av fibrerna i denna plexus representeras av förgrening av dendriterna i de underliggande skikten.
II - yttre granulär skiktet bildas av många små pyramidala och stellate neuroner. Dendriterna av dessa celler stiger in i det molekylära lagret, och axonerna går antingen in vit substans, eller, bildar bågar, kommer också in i tangentiell plexus av fibrer i det molekylära lagret.
III - Det bredaste lagret av hjärnbarken - pyramidal. Den innehåller pyramidala neuroner, Martinotti-celler och spindelceller med en dubbel bukett av dendriter. De apikala dendriterna i pyramiderna går in i det molekylära lagret, de laterala dendriterna bildar synapser med intilliggande celler i detta lager. Pyramidcellens axon avgår alltid från sin bas. I små celler förblir det i cortex, i stora celler bildar det en myelinfiber som går till hjärnans vita substans. Axonerna hos små polygonala Martinotti-celler är riktade mot det molekylära lagret. Pyramidskiktet utför huvudsakligen associativa funktioner.
IV- Inre kornig skiktet i vissa fält av cortex är mycket starkt utvecklat (till exempel i den visuella och auditiva cortex), medan det i andra kan vara nästan frånvarande (till exempel i den precentrala gyrusen). Detta skikt bildas av små stellate neuroner. Den består av ett stort antal horisontella fibrer.
V- Ganglion lager av cortex bildas av stora pyramider, och regionen av den motoriska cortex (precentral gyrus) innehåller jättepyramider , som först beskrevs av Kiev-anatomen V. A. Bets. Pyramidernas apikala dendriter når det första lagret. Pyramidaxoner projekterar till de motoriska kärnorna i hjärnan och ryggrad. De längsta axonerna av Betz-celler i de pyramidala banorna når de kaudala segmenten av ryggmärgen.
Förutom pyramidala neuroner i det ganglioniska lagret av cortex finns det vertikala spindelformade celler, vars axoner stiger i Jag lager cortex och korgceller.
VI - Lager polymorfa celler bildas av neuroner av olika former (fusiform, stellat, Martinotti-celler). Dessa cellers axoner går in i den vita substansen som en del av de efferenta vägarna, och dendriterna når det molekylära lagret.
Myeloarkitektonik
Bland hjärnbarkens nervfibrer kan man urskilja associativ fibrer som förbinder enskilda delar av cortex på en halvklot, kommissural förbinder cortex av olika hemisfärer, och utsprång fibrer, både afferenta och efferenta, som förbinder cortex med kärnorna i de nedre delarna av centrala nervsystemet. Projektionsfibrer i hjärnhalvornas cortex bildar radiella strålar som slutar i det III - pyramidala lagret. Förutom den redan beskrivna tangentiella plexusen i det I - molekylära lagret, på nivån av IV - de inre granulära och V - ganglionlagren, finns det två tangentiella lager av myeliniserade nervfibrer - respektive den yttre remsan av Bayarger och den inre remsa av Bayarger. De två sista systemen är plexusformade av terminalsektionerna av de afferenta fibrerna.
Modulär princip för kortikal organisation
I hjärnhalvornas cortex beskrivs repeterande block, eller moduler, av neuroner, vilka anses vara dess morfofunktionella enheter med förmåga till relativt autonom aktivitet. De har formen av cylindrar, eller kolumner, som passerar vertikalt genom hela tjockleken av cortex.
Varje modul inkluderar afferenta vägar, ett system av lokala förbindelser och efferenta vägar.
TILL afferenta vägar inkluderar kortiko-kortikala och thalamo-kortikala fibrer.
Modulen är organiserad kring kortiko-kortikala fibrer, som är axoner av pyramidceller antingen i samma hemisfär eller i den motsatta. Kortiko-kortikala fibrer bildar ändar i alla lager av cortex i en given modul.
Modulen inkluderar också thalamo-kortikala fibrer som slutar i lager IV av cortex på taggiga stellateuroner och basala dendriter av pyramidala neuroner.
Efferenta vägar bildas av axoner av stora och gigantiska pyramidala neuroner, såväl som axoner av fusiforma och några andra celler i lager VI i cortex.
Lokalt länksystem bildas av modulens interkalära neuroner, som inkluderar mer än ett dussin typer av celler. De flesta av dem är hämmande och reglerar aktiviteten hos övervägande pyramidala neuroner.
Från modulens hämmande neuroner högsta värde ha:
- axo-axonala celler;
- celler - "kandelabre";
- korgceller;
- celler med en dubbel bukett av dendriter;
- celler med en axonal borste.
Systemet av hämmande neuroner spelar rollen som ett filter som hämmar en del av pyramidala nervceller i cortex.
Slidor av hjärnan och ryggmärgen
Hjärnan och ryggmärgen är täckta med tre bindvävsmembran: mjuka, i direkt anslutning till hjärnvävnaderna, arachnoid och solid, som gränsar till benvävnad skalle och ryggrad. Hjärnhinnorna utför ett skyddande, inkl. stötdämpande funktion, ger produktion och absorption av cerebrospinalvätska.
pia mater direkt intill hjärnvävnaden och avgränsad från den av det marginella gliamembranet. i lösa fibrösa bindväv membran finns det ett stort antal blodkärl som matar hjärnan, många nervfibrer, terminalapparat och enstaka nervceller. Pia mater omger kärlen som penetrerar hjärnan och bildar ett perivaskulärt pialmembran runt dem. I hjärnans ventriklar deltar pia mater, tillsammans med ependyma, i bildandet av choroidplexus som producerar cerebrospinalvätska.
Arachnoid representeras av ett tunt lager av lös fibrös bindväv. Mellan den och pia matern ligger ett nätverk av tvärstänger, bestående av tunna buntar av kollagen och tunna elastiska fibrer. Detta nätverk länkar ihop skalen. Mellan pia mater, som upprepar avlastningen av hjärnvävnaden, och arachnoid, som passerar genom de förhöjda områdena utan att gå in i fördjupningarna, finns ett subarachnoid (subaraknoidal) utrymme, genomsyrat av tunt kollagen och elastiska fibrer som förbinder membranen med varandra. Det subarachnoidala utrymmet kommunicerar med hjärnans ventriklar och innehåller cerebrospinalvätska. Detta område är värd för stora blodkärl, vars grenar matar hjärnan.
Villi i arachnoidmembranet (de största kallas pachyonic granulations) fungerar som platser genom vilka ämnen från cerebrospinalvätskan återvänder till blodet. De är avaskulära utväxter av arachnoid, som innehåller ett nätverk av slitsliknande utrymmen och sticker ut i lumen av bihålorna i dura mater.
duralhinnan bildas av tät fibrös bindväv som innehåller många elastiska fibrer. I kranialhålan är den tätt sammansmält med periosteum. I ryggmärgskanalen dura mater avgränsas från kotbenet av ett epiduralt utrymme fyllt med ett lager av lös fibrös bindväv, vilket ger den en viss rörlighet. Mellan dura mater och arachnoid finns det subdurala utrymmet. Det subdurala utrymmet innehåller en liten mängd vätska.
Membranen från sidan av subdural- och subarachnoidutrymmet är täckta med ett lager platta celler glial natur.
Åldersförändringar
Förändringar i centrala nervsystemet i gammal ålder associerade främst med sklerotiska förändringar i cerebrala kärl. I hög ålder tjocknar hjärnans mjuka och arachnoida membran. Kalkavlagringar kan förekomma i dem. Det finns atrofi av cortex halvklot främst i frontal- och parietalloberna. Antalet nervceller per volymenhet av cortex minskar, detta beror främst på celldöd. Neuroner minskar i storlek, förlorar delvis sin basofila substans, kärnorna blir tätare, deras kontur blir ojämn. Pyramiderna i V-skiktet i motorbarken och de päronformade cellerna i cerebellar cortex förändras snabbare än andra. Lipofuscingranulat ackumuleras i nervceller i olika delar av nervsystemet.
Alla de viktigaste mänskliga beteendereaktionerna utförs med hjälp av det centrala nervsystemet. Huvudfunktionerna för CNS är:
- integration av alla delar av kroppen till en enda helhet och deras reglering;
- tillstånd och beteendehantering organism i enlighet med förhållandena yttre miljön och hans behov.
Hos högre djur och människor den ledande delen av CNS är hjärnbarken . Den styr de mest komplexa funktionerna i mänskligt liv mentala processer(medvetande, tänkande, tal, minne, etc.).
De huvudsakliga metoderna för att studera centrala nervsystemets funktioner är metoder för avlägsnande och irritation (på kliniken och på djur), registrering av elektriska fenomen, metoden betingade reflexer.
Nya metoder för att studera det centrala nervsystemet fortsätter att utvecklas: med hjälp av den sk datortomografi du kan se de morfofunktionella förändringarna i hjärnan på olika djup; fotografering i infraröda strålar (termisk avbildning) gör att du kan upptäcka de mest "heta" punkterna i hjärnan; nya data om hjärnans arbete tillhandahålls av studiet av dess magnetiska svängningar.
Grundläggande funktioner och interaktioner av neuroner
De viktigaste strukturella elementen i nervsystemet är nervceller eller neuroner.
Grundläggande funktioner hos neuroner
Genom neuroner överförs information från en del av nervsystemet till en annan, information utbyts mellan nervsystemet och olika delar av kroppen. Neuroner utför komplexa informationsprocesser. Med deras hjälp, svar organism (reflexer) till yttre och inre stimuli.
Således, neuronernas huvudfunktioner är: perception av yttre stimuli receptorfunktion, deras bearbetning integrerande funktion och överföring av nervpåverkan till andra neuroner eller olika arbetsorgan effektorfunktion. I kroppen av nervcellen, eller soma, äger huvudprocesserna för informationsbehandling rum. Många trädgrenade processer dendriter(grekiskt dendronträd) fungerar som neuroningångar genom vilka signaler kommer in i nervcellen. Utgången av en neuron är en process som sträcker sig från cellkroppen axon(grekisk axelaxel), som överför nervimpulser vidare till en annan nervcell eller arbetsorgan (muskel, körtel). Den initiala delen av axonet och expansionen vid platsen för dess utträde från cellkroppen har en särskilt hög excitabilitet axon kulle nervcell. Det är i detta segment av cellen som nervimpulsen uppstår.
Typer av neuroner
Neuroner delas in i tre huvudtyper: afferenta, efferenta och intermediära. Afferenta neuroner(känslig eller centripetal) överför information från receptorer till 1CNS. Dessa nervcellers kroppar är belägna utanför CNS i spinal noder och i noder kranialnerver. Afferenta neuroner har en lång process - en dendrit, som kommer i kontakt i periferin med en uppfattande formation - en receptor eller bildar en receptor själv, samt en andra process - ett axon som går in genom bakre horn in i ryggmärgen.
Efferenta neuroner(centrifugal) är förknippade med överföringen av nedåtgående influenser från de överliggande golven i nervsystemet till de underliggande eller från det centrala nervsystemet till de arbetande organen. Efferenta neuroner kännetecknas av ett förgrenat nätverk av korta processer - dendriter och en lång process - axon.
mellanliggande neuroner(interneuroner, eller intercalary) - dessa är som regel mindre celler som kommunicerar mellan olika (särskilt afferenta och efferenta) neuroner. De överför nervpåverkan i horisontell riktning (till exempel inom ett segment av ryggmärgen) och i vertikal riktning (till exempel från ett segment av ryggmärgen till andra - högre eller lägre segment). På grund av de många grenarna av axonet kan mellanliggande neuroner samtidigt excitera ett stort antal andra neuroner.
Excitatoriska och hämmande synapser
Interaktionen mellan neuroner med varandra (och med effektororgan) sker genom specialundervisning synapser (grekisk kontakt). De bildas av terminala grenar av en neuron på kroppen eller processer av en annan neuron. Ju fler synapser på en nervcell, desto mer uppfattar den olika stimuli och, följaktligen, desto bredare är inflytandesfären på dess aktivitet och möjligheten att delta i olika reaktioner i kroppen. Det finns särskilt många synapser i de högre delarna av nervsystemet, och det finns i neuroner med de mest komplexa funktionerna.
Tre element urskiljs i synapsstrukturen (Fig. 1):
- presynaptiskt membran, bildad av en förtjockning av membranet i axonets terminala gren;
- synaptisk klyfta mellan neuroner;
- postsynaptisk membran förtjockning av den intilliggande ytan av nästa neuron.
Ris. 1. Diagram över en synaps
Pre. presynaptiskt membran,
Snabb. postsynaptisk membran,
C synoptiska bubblor,
Щ synoptisk gap,
M mitokondrier,
Ah acetylkolin
P-receptorer och porer (porer)
dendrit (D) nästa
nervcell.
Pil ensidig ledning av excitation.
I de flesta fall utförs överföringen av påverkan från en neuron till en annan kemiskt. Den presynaptiska delen av kontakten innehåller synaptiska vesiklar som innehåller speciella ämnen medlare eller mellanhänder. De kan vara acetylkolin (i vissa celler i ryggmärgen, i autonoma noder), noradrenalin (i ändarna av sympatiska nervfibrer, i hypotalamus), vissa aminosyror Nervimpulser som anländer till slutet av axonet orsakar tömning av de synaptiska vesiklerna och utsöndringen av mediatorn i den synaptiska klyftan.
Genom arten av påverkan på den efterföljande nervcellen särskiljs excitatoriska och hämmande synapser.
Vid excitatoriska synapser mediatorer (till exempel acetylkolin) binder till specifika makromolekyler i det postsynaptiska membranet och orsakar dess depolarisering. I detta fall en liten och kortvarig (ca 1 ms) fluktuation av membranpotentialen mot avpolarisering eller excitatorisk postsynaptisk potential(VPSP). För att excitera en neuron är det nödvändigt för EPSP att nå en tröskelnivå. För detta måste storleken på depolarisationsförskjutningen av membranpotentialen vara minst 10 mV. Medlarens verkan är mycket kort (1-2 ms), varefter den delas upp i ineffektiva komponenter (till exempel, acetylkolin bryts ned av enzymet kolinesteras till kolin och ättiksyra ) silt och återabsorberas tillbaka av presynaptiska ändar (till exempel noradrenalin).
I hämmande synapser innehöll bromsval (Till exempel, gamma-aminosmörsyra ). Deras verkan på det postsynaptiska membranet orsakar en ökning av frisättningen av kaliumjoner från cellen och en ökning av polariseringen av membranet. I detta fall registreras en kortvarig fluktuation av membranpotentialen i riktningen hyperpolarisering hämmande postsynaptisk potential(TPSP). Som ett resultat hämmas nervcellen. Det är svårare att excitera det än i initialtillståndet. Detta kommer att kräva en starkare stimulering för att nå en kritisk nivå av depolarisering.
Uppkomsten av ett impulssvar från en neuron
Både excitatoriska och hämmande synapser finns på kroppens membran och dendriter i nervcellen. . Vid vissa tidpunkter kan vissa av dem vara inaktiva, medan den andra delen har en aktiv effekt på membranområdena intill dem. Den övergripande förändringen i membranpotentialen hos en neuron är resultatet av komplex interaktion(integration) av lokala EPSP:er och IPSP:er av alla många aktiverade synapser. Med samtidig påverkan av både excitatoriska och hämmande synapser sker en algebraisk summering (dvs ömsesidig subtraktion) av deras effekter. I det här fallet kommer excitationen av neuronen endast att ske om summan av excitatoriska postsynaptiska potentialer blir mer än beloppet hämmande . Detta överskott bör vara ett visst tröskelvärde (cirka 10 mV). Endast i detta fall uppträder cellens aktionspotential. Det bör noteras att i allmänhet beror excitabiliteten hos en neuron på dess storlek: ju mindre cellen är, desto högre är dess excitabilitet .
Med uppkomsten av en handlingspotential, processen att genomföra nervimpuls längs axonet och överför det till nästa neuron eller arbetsorgan, dvs. neurons effektorfunktion utförs. Nervimpulsen är det huvudsakliga kommunikationsmedlet mellan neuroner.
Således, överföring av information i nervsystemet sker genom två elektriska mekanismer (EPSP; IPSP; handlingspotential) och kemiska (förmedlare).
Funktioner av aktiviteten hos nervcentra
Egenskaperna hos nervcentra är till stor del relaterade till egenskaperna hos ledningen av nervimpulser genom synapser som förbinder olika nervceller.
Funktioner av ledning av excitation genom nervcentra
nervcentrum kallas helheten av nervceller som är nödvändiga för genomförandet av någon funktion. Dessa centra svarar med lämpliga reflexreaktioner på extern stimulering från de receptorer som är associerade med dem. Cellerna i nervcentra reagerar också på sin direkta irritation av ämnen i blodet som strömmar genom dem (humorala influenser). I hela organismen finns en strikt koordination - samordning av sin verksamhet.
Ledningen av en excitationsvåg från en neuron till en annan genom synapsen sker i de flesta nervceller på kemisk väg med hjälp av en mediator, och mediatorn finns endast i den presynaptiska delen av synapsen och saknas i det postsynaptiska membranet. Det är därför viktig funktion bedriva excitation genom synoptiska kontakter är ensidigt innehav nervpåverkan, vilket är möjligt endast från det presynaptiska membranet till det postsynaptiska och är omöjligt i motsatt riktning. I detta avseende har flödet av nervimpulser i reflexbågen en viss riktning från afferenta neuroner till interkalär och sedan till efferenta motoneuroner eller autonoma neuroner.
Stor betydelse i nervsystemets aktivitet har en annan egenskap att leda excitation genom synapser långsam ledning. Den tid som spenderas på de processer som sker från det att nervimpulsen närmar sig det presynaptiska membranet tills potentialer uppstår i det postsynaptiska membranet kallas synaptisk fördröjning. I de flesta centrala neuroner är det cirka 0,3 ms. Efter detta krävs mer tid för utvecklingen av den excitatoriska postsynaptiska potentialen (EPSP) och aktionspotentialen. Hela processen med nervimpulsöverföring (från en cells aktionspotential till nästa cells aktionspotential) genom en synaps tar ungefär 1,5 ms. Med trötthet, nedkylning och ett antal andra influenser ökar varaktigheten av den synaptiska fördröjningen. Om det krävs deltagande för genomförandet av någon reaktion ett stort antal neuroner (många hundra och till och med tusentals), då kan den totala fördröjningen i ledning genom nervcentra vara tiondels sekund och till och med hela sekunder.
Med reflexaktivitet total tid från det ögonblick som en extern stimulans appliceras på utseendet av ett svar från kroppen - den så kallade latent eller latent reflextid bestäms huvudsakligen av varaktigheten av ledning genom synapserna. Värdet på den latenta tiden för reflexen fungerar som en viktig indikator funktionellt tillstånd nervcentra . Mätning av den latenta tiden för ett enkelt mänskligt motoriskt svar på en extern signal används ofta i praktiken för att bedöma det funktionella tillståndet hos CNS (Fig. 2).
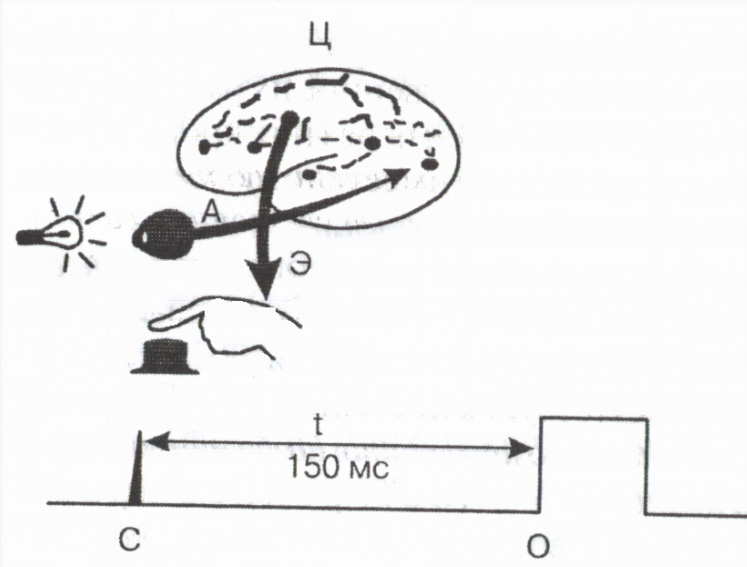
Ris. 2. Schema för att mäta tiden för motorisk reaktion
En afferent
E efferenta och C centrala vägar; C - ljussignalmärke,
О-knappen tryck på markeringen,
t 150ms svarstid.
Excitation Summation
Som svar på en enda afferent våg från receptorer till neuroner frigörs en liten mängd mediator i den presynaptiska delen av synapsen. I det här fallet uppstår EPSP vanligtvis i neurons postsynaptiska membran - en liten lokal depolarisering. För att det totala EPSP-värdet över hela neuronmembranet ska nå tröskeln för uppkomsten av en aktionspotential krävs summering av många subtröskel-EPSP på cellmembranet. Endast som ett resultat av en sådan summering av excitation svarar en neuron. Skilj mellan rumslig och tidsmässig summering.
Rumslig summering observeras vid samtidig mottagning av flera impulser i samma neuron genom olika presynaptiska fibrer. Samtidig excitation av synapser i olika delar av neuronmembranet ökar amplituden av den totala EPSP till ett tröskelvärde. Som ett resultat uppstår en svarsimpuls från neuronen och en reflexreaktion utförs. Till exempel kräver att erhålla ett ryggmärgsmotorcellsvar vanligtvis samtidig aktivering av 50-100 afferenta fibrer från motsvarande perifera receptorer.
Tidssumma inträffar när samma afferenta väg aktiveras av en serie på varandra följande stimuli. Om intervallen mellan inkommande impulser är tillräckligt korta, hinner inte neurons EPSP från tidigare stimuli blekna, sedan överlagras efterföljande EPSP på varandra tills depolariseringen av neuronmembranet når en kritisk nivå för uppkomsten av en åtgärd potential. På det här sättet till och med svaga irritationer efter en tid kan de orsaka reaktioner från kroppen (till exempel nysningar och hosta som svar på mild irritation av slemhinnan i luftvägarna).
Transformation och assimilering av rytm
Arten av svarsurladdningen från en neuron beror inte bara på stimulans egenskaper, utan också på det funktionella tillståndet hos själva neuronen.(dess membranladdning, excitabilitet, labilitet). Nervceller har förmågan att ändra frekvensen av överförda impulser, d.v.s. rytmomvandlingsegenskap.
Med hög excitabilitet hos en neuron (till exempel efter att ha tagit koffein), ökad impulsering (rytmmultiplikation) och med låg excitabilitet (till exempel med trötthet) rytmnedgång , eftersom flera inkommande impulser måste summeras för att slutligen nå tröskeln för uppkomsten av en aktionspotential. Dessa förändringar i frekvensen av impulser kan förstärka eller försvaga kroppens svar på yttre stimuli.
Med rytmiska stimuli kan aktiviteten hos en neuron stämma in på rytmen av inkommande impulser, dvs. rytminlärningsfenomen(Ukhtomsky A.A., 1928). Utvecklingen av rytm assimilering ger inställning av aktiviteten hos många nervcentra när man hanterar komplexa motoriska handlingar är det särskilt viktigt att bibehålla tempot i cykliska övningar.
spåra processer
Efter slutet av stimulans verkan fortsätter det aktiva tillståndet hos nervcellen eller nervcentret vanligtvis under en tid. Varaktigheten av spårprocesser är olika: kort i ryggmärgen (flera sekunder eller minuter), mycket längre i hjärnans centrum (tiotals minuter, timmar eller till och med dagar) och mycket lång i hjärnbarken (upp till flera decennier). ).
Impulser som cirkulerar genom slutna kretsar av neuroner kan upprätthålla ett tydligt och kortvarigt tillstånd av excitation i nervcentrum. Långsiktiga dolda spår är mycket svårare i naturen. Det antas att långsiktigt bevarande av spår i nervcellen med alla karakteristiska egenskaper stimulans är baserad på en förändring i strukturen hos de proteiner som utgör cellen och på omstruktureringen av synaptiska kontakter.
Korta impulsefterverkningar (varar upp till 1 timme) ligger till grund för den sk korttidsminne , och långvariga spår associerade med strukturella och biokemiska omarrangemang i celler, är grunden för bildningen långtids minne.
CNS-koordination
Processerna för koordinering av aktiviteten i det centrala nervsystemet är baserade på koordineringen av två nervprocesser - excitation och hämning. Hämning är en aktiv nervös process , som förhindrar eller hämmar upphetsning.
Värdet av hämningsprocessen i centrala nervsystemet
Fenomenet med hämning i nervcentra upptäcktes först av I. M. Sechenov 1862. Betydelsen av denna process övervägdes av honom i boken Reflexes of the Brain (1863).
Genom att doppa grodbenet i syra och samtidigt irritera vissa delar av hjärnan (till exempel genom att applicera en kristall bordssalt på diencephalonområdet) observerade I. M. Sechenov en kraftig fördröjning och till och med fullständig frånvaro av ryggmärgens "sura" reflex (utdragning av tass). Därför drog han slutsatsen att vissa nervcentra avsevärt kan förändra reflexaktiviteten i andra centra, i synnerhet kan de överliggande nervcentra hämma aktiviteten hos de underliggande. Den beskrivna upplevelsen kom in i fysiologins historia under namnet Sechenov hämning .
Hämmande processer är en nödvändig komponent i samordningen av nervös aktivitet. För det första, hämningsprocessen begränsar spridningen av excitation till närliggande nervcentra, vilket bidrar till dess koncentration i de nödvändiga delarna av nervsystemet. För det andra, uppstår i vissa nervcentra parallellt med exciteringen av andra nervcentra, inhiberingsprocessen därigenom stänger av onödiga aktiviteter det här ögonblicket kroppar . För det tredje skyddar utvecklingen av hämning i nervcentra dem från överdriven överbelastning under arbetet, det vill säga det spelar en skyddande roll.
Postsynaptisk och presynaptisk hämning
Bromsprocess , i motsats till excitation, kan inte fortplanta sig längs nervfibern det är alltid en lokal process inom området synaptiska kontakter. Beroende på ursprungsplatsen särskiljs presynaptisk och postsynaptisk hämning.
Postsynaptisk hämning dessa är hämmande effekter som förekommer i det postsynaptiska membranet. Oftast är denna typ av hämning associerad med närvaron i det centrala nervsystemet av speciell hämmande neuroner. De är en speciell typ av interkalära neuroner, där axonändar frisätter en hämmande neurotransmittor. En av dessa medlare är gamma-aminosmörsyra (GAM K).
Nervimpulser som närmar sig hämmande neuroner orsakar samma excitationsprocess i dem som i andra nervceller. Som svar fortplantar sig den vanliga aktionspotentialen längs den hämmande cellens axon. Men till skillnad från andra neuroner, släpper axonändarna inte en excitatorisk, utan en hämmande signalsubstans. Som ett resultat hämmar hämmande celler de neuroner på vilka deras axoner slutar.
Särskilda hämmande neuroner inkluderar Ranshaw-celler i ryggmärgen, Purkinje-celler i cerebellum, korgceller i diencephalon, etc. Till exempel är hämmande celler av stor betydelse för att reglera aktiviteten hos antagonistmuskler: genom att leda till avslappning av antagonistmusklerna underlättar de därigenom den samtidiga sammandragningen av agonistmusklerna (fig. 3).
Renshaw celler delta i regleringen av aktivitetsnivån hos enskilda motorneuroner i ryggmärgen. När en motorneuron är exciterad, färdas impulser längs dess axon till muskelfibrerna och samtidigt längs axonens kollateraler till Ranshaw-hämmande cell. Den senares axoner "återgår" till samma neuron, vilket orsakar dess hämning. Ju fler excitatoriska impulser motorneuronen skickar till periferin (och därmed till den hämmande cellen), desto starkare är detta återbromsning(ett slags postsynaptisk hämning). Sådan slutet system fungerar som en självreglerande mekanism för neuronen och skyddar den från överdriven aktivitet.
Purkinjeceller i lillhjärnan deras hämmande effekter på cellerna i de subkortikala kärnorna och stamstrukturerna är involverade i regleringen av muskeltonus.
korgceller i diencephalon är de så att säga grindar som tillåter eller inte skickar impulser som går till hjärnbarken från olika delar av kroppen.
presynaptisk hämning sker före synaptisk kontakt i den presynadtiska regionen. Änden av den hämmande nervcellens axon bildar en synaps i slutet av den excitatoriska nervcellens axon, vilket orsakar en alltför kraftig depolarisering av membranet i denna axon, vilket hämmar de aktionspotentialer som passerar hit och därigenom blockerar överföringen av excitation. Denna typ av hämning begränsar flödet av afferenta impulser till nervcentra, vilket stänger av influenser som är främmande för huvudaktiviteten.
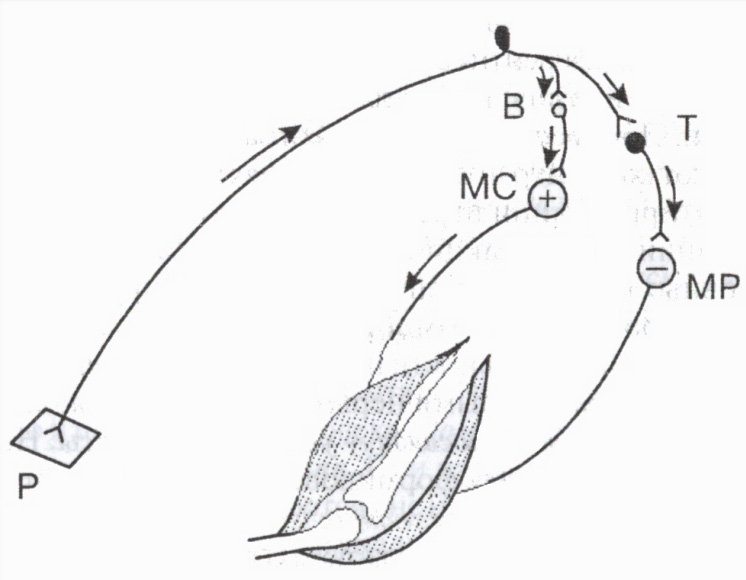
Ris. 3. Inblandning av den hämmande cellen i regleringen av antagonistmuskler
B och T - excitatoriska och hämmande neuroner. Excitation (+) motorneuron av flexormuskeln (MS) och inhibering (-) motorneuron av extensormuskeln (MR). P hudreceptor.
Fenomenet bestrålning och koncentration
När en receptor stimuleras kan excitation i princip spridas i CNS i valfri riktning och till vilken nervcell som helst. Detta beror på de många sammankopplingarna av neuroner hos en reflexbåge med neuroner av andra reflexbågar. Spridningen av excitationsprocessen till andra nervcentra kallas fenomenet bestrålning.
Ju starkare afferent stimulering och ju högre excitabilitet hos de omgivande neuronerna, desto fler neuroner täcks av bestrålningsprocessen. Inhiberingsprocesserna begränsar bestrålningen och bidrar till koncentrationen av excitation vid startpunkten av CNS.
Bestrålningsprocessen spelar en viktig roll positiv roll i bildandet av nya reaktioner av kroppen (orienterande reaktioner, betingade reflexer). Ju mer olika nervcentra aktiveras, desto lättare är det att välja bland dem de centra som behövs mest för efterföljande aktivitet. Tack vare bestrålning excitationer mellan olika nervcentra nya funktionella relationer uppstår betingade reflexer . Utifrån detta är det till exempel möjligt att forma nya motoriska färdigheter.
Samtidigt kan bestrålning av excitation också negativ påverkan på kroppens tillstånd och beteende, vilket stör de subtila relationerna mellan upphetsade och hämmade nervcentra och orsakar rörelsekoordinationsstörningar .
Dominerande
Genom att undersöka egenskaperna hos intercentrala relationer upptäckte A. A. Ukhtomsky att om en komplex reflexreaktion inträffar i djurkroppen, till exempel upprepade sväljningshandlingar, så slutar den elektriska stimuleringen av motorcentra inte bara att orsaka rörelse av extremiteterna i detta ögonblick , men ökar också flödet av kedjereaktionen som har börjat , svälja, som visade sig vara dominerande.
Ett sådant dominerande excitationsfokus i det centrala nervsystemet, som bestämmer organismens nuvarande aktivitet, betecknades av AA Ukhtomsky (1.923) med termen dominant.
Dominant fokus kan uppstå när förhöjd nivå excitabilitet av nervceller, som skapas av olika humorala och nervösa influenser. Det undertrycker aktiviteten hos andra centra, vilket ger konjugerad hämning.
Enandet av ett stort antal neuroner till ett dominant system sker genom ömsesidig anpassning till den allmänna aktivitetstakten, d.v.s. genom assimileringen av rytmen. Vissa nervceller bromsar sin högre aktivitetshastighet, medan andra ökar sin låga hastighet till en genomsnittlig, optimal rytm. Den dominanta kan förbli under lång tid i ett dolt, spårtillstånd (potentiellt dominant). När den tidigare staten eller den tidigare yttre situationen återupptas kan den dominerande återuppstå (aktualisering av den dominerande). Till exempel, i pre-launch-tillståndet, aktiveras alla de nervcentra som var en del av arbetssystemet under tidigare träningar, och följaktligen förbättras funktionerna i samband med arbete. mental avrättning träning eller representationen av rörelser reproducerar också den arbetande dominanten, vilket ger träningseffekten av representationen av rörelser och är grunden för den så kallade ideomotoriska träningen. När du är helt avslappnad (t.ex. när autogen träning) idrottare uppnår eliminering av arbetande dominanter, vilket påskyndar återhämtningsprocessen.
Som en beteendefaktor förknippas dominant med högre nervös aktivitet och mänsklig psykologi. Dominant är fysiologisk grund uppmärksamhetshandling. I närvaro av en dominant förblir många influenser från den yttre miljön utanför vår uppmärksamhet, men de som är av särskilt intresse för oss fångas och analyseras mer intensivt. Sålunda är den dominanta en kraftfull faktor i urvalet av biologiskt och socialt de viktigaste stimulierna.
Funktioner i ryggmärgen och subkortikala regioner i hjärnan
I CNS, äldre segmentell och evolutionärt yngre suprasegmentell delar av nervsystemet. De segmentella sektionerna inkluderar ryggraden, medulla oblongata och mellanhjärnan, vars sektioner reglerar funktionerna hos enskilda delar av kroppen som ligger på samma nivå. Suprasegmental divisioner diencephalon, cerebellum och cerebral cortex har inte direkta kopplingar till kroppens organ, utan styr sin aktivitet genom de underliggande segmental divisionerna.
Ryggrad
Ryggmärgen är den lägsta och äldsta delen av CNS. . Den grå substansen i den mänskliga ryggmärgen innehåller cirka 13,5 miljoner nervceller. Av dem huvuddelen (97 %) är mellanliggande celler (interneuroner eller interneuroner) , som tillhandahåller komplexa koordinationsprocesser inom ryggmärgen. Bland de motoriska nervcellerna i ryggmärgen, stora alfamotoriska neuroner och liten gammamotoriska neuroner. De tjockaste och snabbast ledande fibrerna i motornerverna avgår från alfamotorneuronerna, orsakar förkortningar skelett- muskelfibrer. Tunna fibrer av gammamotorneuroner orsakar inte muskelkontraktion. De passar pro-prioreceptorerna muskelspindlar och justera deras känslighet.
Ryggmärgsreflexer kan delas in i motor utförs av alfamotoriska neuroner i de främre hornen, och vegetativ utförs av afferenta celler i sidohornen.
Motorneuroner i ryggmärgen innerverar alla skelettmuskler (med undantag för ansiktsmusklerna) . Ryggmärgen bär ut elementära motoriska reflexer böjning och extensor, rytmisk, stegande, som härrör från irritation av huden eller proprioreceptorer av muskler och senor, och skickar också konstanta impulser till musklerna, stödjande muskeltonus. Särskilda motorneuroner innerverar andningsmusklerna - interkostalmusklerna och diafragman, och ger andningsrörelser. Autonoma neuroner innerverar alla inre organ (hjärta, blodkärl, svettkörtlar, körtlar inre sekretion, matsmältningskanalen, urinvägarna).
Ledarfunktion av ryggmärgen är förknippad med överföringen till de överliggande delarna av nervsystemet av flödet av information som tas emot från periferin och med ledning av impulser som kommer från hjärnan till ryggmärgen.
Bakom senaste åren tagit fram speciella tekniker att studera ryggmärgens aktivitet i frisk person. Exempelvis bedöms alfamotorneuronernas funktionstillstånd genom förändringar i musklernas responspotentialer vid perifera stimuli, s.k. H-reflex (Hoffmann reflex) vadmuskel när man är irriterad tibialisnerven och genom att T-reflex (från senan) soleusmuskel med irritation av akillessenan. Tekniker har utvecklats för att registrera (från intakta integument i kroppen) potentialer som passerar genom ryggmärgen till hjärnan.
medulla oblongata och pons varolii
Medulla oblongata och pons (i allmänhet, bakhjärnan) är en del av hjärnstammen. Här finns en stor grupp kranialnerver (från V till XII par), som innerverar hud, slemhinnor, muskler i huvudet och ett antal inre organ(hjärta, lungor, lever). Här finns centra för många matsmältningsreflexer tugga, svälja, rörelser i magen och delar av tarmarna, utsöndring av matsmältningssafter och centra för vissa skyddsreflexer (nysningar, hosta, blinkande, tårar, kräkningar) och centra för vatten-salt- och sockermetabolism . I botten av IV ventrikeln i medulla oblongata finns ett vitalt andningscentrum, bestående av centra för inandning och utandning. Den består av små celler som skickar impulser till andningsmusklerna genom ryggmärgens motorneuroner.
Ligger i omedelbar närhet kardiovaskulärt centrum . Dess stora celler reglerar aktiviteten i hjärtat och blodkärlens lumen. Sammanflätningen av cellerna i andnings- och kardiovaskulära centra säkerställer deras nära interaktion.
Medulla oblongata spelar en viktig roll i genomförandet av motoriska handlingar och i regleringen av skelettmuskeltonus, öka tonen i sträckmusklerna . Han deltar i synnerhet i genomförandet posturala reflexer (cervikal, labyrintisk). Stigande passerar genom medulla oblongata auditiva, vestibulära, proprioceptiva och taktila vägar .
mellanhjärnan
Mellanhjärnan innehåller quadrigemina, substantia nigra och röda kärnor. I de främre tuberklerna i quadrigemina finns visuella subkortikala centra , och på baksidan auditiv . Mellanhjärnan är involverad i reglering av ögonrörelser , Utför pupillreflex (pupillutvidgning i mörker och förträngning i ljus).
Quadrigemina utför en serie reaktioner som är komponenter i orienteringsreflexen . Som svar på en plötslig irritation vänder sig huvudet och ögonen mot stimulansen, och hos djur är öronen alerta. Denna reflex (enligt I. P. Pavlov, "Vad är det?"-reflexen) är nödvändig för att förbereda kroppen för ett snabbt svar på varje ny påverkan.
svart substans mellanhjärnan är relaterad till reflexerna av att tugga och svälja, är involverad i reglering av muskeltonus (speciellt när man gör det små rörelser fingrar) och i organisationen av vänliga motoriska reaktioner.
röd kärna mellanhjärnan utför motoriska funktioner reglerar skelettmuskeltonus , vilket orsakar en ökning av tonen i flexormusklerna. Med en betydande effekt på skelettmuskeltonus är mellanhjärnan involverad i ett antal justering av reflexer för att bibehålla hållningen (upprätta inställning av kroppen med kronan uppåt, etc.).
diencephalon
Diencephalon består av thalamus (syntuberkler) och hypotalamus (hypothalamus).
Genom talamus passera genom alla afferenta vägar (med undantag för de olfaktoriska), som skickas till motsvarande uppfattande områden i cortex (auditiva, visuella, etc.). Thalamus kärnor är indelade i specifika och ospecifika . De specifika är switchande (relä) kärnor och associativa . Genom thalamus kopplingskärnor överförs afferenta influenser från alla receptorer i kroppen. Associativa kärnor tar emot impulser från kopplingskärnorna och säkerställer deras interaktion. Utöver dessa kärnor innehåller thalamus ospecifika kärnor som har både aktiverande och hämmande effekter på små områden av cortex.
På grund av omfattande kopplingar spelar thalamus en avgörande roll i kroppens liv. Impulser som kommer från thalamus till cortex förändrar tillståndet hos kortikala neuroner och reglerar rytm av kortikal aktivitet . Med direkt deltagande av thalamus sker bildandet av betingade reflexer och utvecklingen av motorik, bildandet av känslor man, hans ansiktsuttryck. Talamus spelar en viktig roll i genereringen av förnimmelser, i synnerhet förnimmelser av smärta . Förknippad med hans arbete reglering av biorytmer i mänskligt liv (dagligt, säsongsbetonat, etc.).
Hypotalamus är det högsta subkortikala regleringscentret autonoma funktioner , stater vakenhet och sömn . Här är de vegetativa centra, reglerar ämnesomsättningen och kroppen, tillhandahållande bibehålla en konstant kroppstemperatur (i varmblodiga) och normal nivå blodtryck stödjande vattenbalans, reglerar känslor av hunger och mättnad . Irritation av de bakre kärnorna i hypotalamus orsakar en ökning av sympatiska influenser och främre - parasympatiska effekter.
På grund av kopplingen mellan hypotalamus och hypofysen ( hypotalamus-hypofyssystemet) aktiviteten hos de endokrina körtlarna kontrolleras. Vegetativa och hormonella svar som regleras av hypotalamus är komponenter i mänskliga känslomässiga och motoriska reaktioner.
Ospecifikt hjärnsystem
Ospecifikt system upptar den mellersta delen av hjärnstammen. Det är inte kopplat till analysen av någon specifik känslighet eller med utförandet av vissa reflexreaktioner. Impulser kommer in i detta system genom laterala förgreningar från alla specifika vägar, som ett resultat av vilket deras omfattande interaktion säkerställs. För icke-specifikt system platsen för neuroner i form av ett diffust nätverk, överflöd och mångfald av deras processer är karakteristiska. Av denna anledning fick hon namnet retikulär bildning eller retikulär bildning.
Skilja på två typer av påverkan icke-specifikt system till arbetet i andra nervcentra aktiverande och hämmande . Båda typerna av dessa influenser kan vara stigande (till de överliggande centran) och fallande (till de underliggande centran). De tjänar för reglering av hjärnans funktionella tillstånd, nivån av vakenhet och reglering av postural-toniska och fasiska reaktioner av skelettmuskler.
Lilla hjärnan
Lillhjärnan är en suprasegmentell formation som inte har direkta kopplingar till den verkställande apparaten. Lillhjärnan består av en oparad formation - vermis och parade halvklot.
De viktigaste nervcellerna i cerebellar cortex är många Purkinje-celler. Tack vare omfattande anslutningar (upp till 200 000 synapser slutar på varje cell), integration av en mängd olika sensoriska influenser , främst proprioceptiva, taktila och vestibulära. Representationen av olika perifera receptorer i cerebellar cortex har somatotop organisation (grekiska somatos kropp, topos plats), det vill säga återspeglar ordningen för deras placering i människokroppen. Dessutom motsvarar detta arrangemang samma arrangemang av representationen av kroppsdelar i hjärnbarken, vilket underlättar informationsutbytet mellan cortex och cerebellum och säkerställer deras gemensamma aktivitet för att kontrollera mänskligt beteende. Den korrekta geometriska organisationen av cerebellära neuroner bestämmer dess vikt vid nedräkning och noggrant underhåll av takten i cykliska rörelser.
Lillhjärnans huvudfunktion är regleringen av posturala toniska reaktioner och koordineringen av motorisk aktivitet.(Orbeli L.A., 1926).
Förbi anatomiska egenskaper(kopplingar av cerebellar cortex med dess kärnor) och lillhjärnans funktionella betydelse delas in i tre längsgående zoner:
- inre eller mediala bark av masken, vars funktion är regleringen av skelettmuskeltonen, upprätthåller kroppens hållning och balans;
- mellanliggande mittdelen hjärnbarken lillhjärnan, vars funktion är att koordinera posturala reaktioner med rörelser och korrigera fel;
- lateral eller lateral cerebral cortex lillhjärnan, som tillsammans med diencephalon och hjärnbarken är involverad i programmeringen av snabba ballistiska rörelser (kast, slag, hopp, etc.).
Basala kärnor
De basala ganglierna inkluderar striatum, som består av caudatkärnan och skalet, och den bleka kärnan, och anses för närvarande också amygdala(relatera till vegetativa centra limbiska systemet) och substantia nigra i mellanhjärnan.
Afferenta influenser kommer till basalkärnorna från kroppens receptorer genom thalamus och från alla områden i hjärnbarken. De kommer nästan uteslutande in i striatum. Efferenta influenser från den skickas till den bleka kärnan och vidare till det extrapyramidala systemets stamcentrum, och även genom thalamus tillbaka till cortex.
De basala kärnorna är involverade i bildandet av betingade reflexer och implementeringen av komplex obetingade reflexer(defensiv, livsmedelsproducerande, etc.). De ger kroppens nödvändiga position under fysiskt arbete, såväl som flödet av automatiska rytmiska rörelser (gamla automatismer).
Den bleka kärnan utför den huvudsakliga motorfunktionen, och striatum reglerar dess aktivitet. För närvarande har betydelsen av kaudatkärnan i kontrollen av komplexa mentala processer - uppmärksamhet, minne, feldetektering - avslöjats.
Alla kroppsfunktioner kan villkorligt delas in i somatiska, eller djur(djur) förknippade med perception extern information och aktiviteten hos muskler, och vegetativ (vegetativ) associerad med aktiviteten hos inre organ, andningsprocesser, blodcirkulation, matsmältning, utsöndring, metabolism, tillväxt och reproduktion.
Funktionell organisation av det autonoma nervsystemet
autonoma nervsystemet kallas en uppsättning efferenta nervceller i ryggmärgen och hjärnan, samt celler av speciella noder (ganglier) som innerverar inre organ. Irritationer av olika receptorer i kroppen kan orsaka förändringar i både somatiska och autonoma funktioner, eftersom de afferenta och centrala delarna av dessa reflexbågar är vanliga. De skiljer sig bara åt i sina olika indelningar. karaktäristiskt drag efferenta banor som ingår i reflexbågarna hos vegetativa reflexer är deras två-neuronstruktur(en neuron finns i det centrala nervsystemet, den andra är i ganglierna eller i det innerverade organet).
Vegetativ nervsystem uppdelad i två divisioner sympatisk och parasympatisk (Fig. 4).
Efferenta vägar sympatiska nervsystemet börjar i bröstet ländryggen ryggmärgen från nervcellerna i dess laterala horn. Överföringen av excitation från de prenodala sympatiska fibrerna till de postnodala sker under medverkan av mediatorn acetylkolin , och från postnodala fibrer till innerverade organ med deltagande av en mediator noradrenalin . Undantagen är fibrerna som innerverar svettkörtlarna och vidgar skelettmuskulaturens kärl, där excitation överförs med acetylkolin.
Efferenta vägar parasympatiska nervsystemet börja i hjärnan från några kärnor i mitten och förlängda märgen, och i ryggmärgen från neuroner sakrala avdelningen. Ledning av excitation i synapserna i den parasympatiska vägen sker med medlarens deltagande acetylkolin . Den andra efferenta neuronen är belägen i eller nära det innerverade organet.
Den högsta regulatorn av autonoma funktioner är hypotalamus. , som verkar i samband med den retikulära bildningen och det limbiska systemet under kontroll av hjärnbarken. Dessutom neuroner som finns i själva organen eller i sympatiska noder, kan utföra sina egna reflexreaktioner utan deltagande av det centrala nervsystemet "perifera reflexer" .
Ris. 4. Autonoma nervsystemet
Vänster fiberutgångsområde: parasympatisk (svart)
och sympatiska (skuggade) system.
Till höger strukturen av den efferenta delen av reflexbågen av vegetativa reflexer. Till vänster är ett diagram över mitten, märgen och ryggmärgen.
Arabiska siffror antal bröstkorgssegment, romerska tal för ländryggssegment.
Det sympatiska nervsystemets funktioner
Med deltagande av det sympatiska nervsystemet uppstår många viktiga reflexer i kroppen, som syftar till att säkerställa dess aktiva tillstånd, inklusive dess motoriska aktivitet. Dessa inkluderar reflexer utvidgning av bronkerna, ökad och ökad hjärtfrekvens , expansion av kärlen i hjärtat och lungorna med samtidig förträngning av kärlen i huden och organen bukhålan (säkerställa omfördelning av blod ), frisättning av avsatt blod från levern och mjälten, nedbrytningen av glykogen till glukos i levern ( mobilisering av kolhydratenergikällor ), ökad aktivitet hos de endokrina körtlarna och svettkörtlarna. Sympatiskt nervsystem minskar aktiviteten hos ett antal inre organ: som ett resultat av vasokonstriktion i njurarna minskar urineringsprocesserna, organens sekretoriska och motoriska aktivitet hämmas mag-tarmkanalen; urinering förhindras väggmuskeln slappnar av Blåsa och dess sfinkter reduceras.
Ökad aktivitet i kroppen åtföljs av sympatisk pupillvidgningsreflex . Av stor betydelse för kroppens motoriska aktivitet är trofisk påverkan av sympatiska nerver på skelettmuskler , förbättra deras ämnesomsättning och funktionella tillstånd, lindra trötthet.
Den sympatiska uppdelningen av nervsystemet ökar inte bara kroppens funktionsnivå utan också mobiliserar sina dolda funktionella reserver , aktiverar hjärnaktivitet, ökar skyddsreaktioner ( immunreaktioner, barriärmekanismer etc.), utlöser hormonella reaktioner. Det sympatiska nervsystemet är av särskild betydelse. i utvecklingen stressiga förhållanden , i de flesta svåra förhållanden livsviktig aktivitet. L. A. Orbeli betonade grundläggande sympatiska influenser för anpassning (anpassning) av kroppen till hårt arbete, till olika förutsättningar yttre miljön. Denna funktion fick namnet adaptiv-trofisk.
Det parasympatiska nervsystemets funktioner
parasympatiska nervsystemet Utför förträngning av bronkerna, bromsning och försvagning av hjärtsammandragningar: sammandragning av hjärtats kärl; påfyllning av energiresurser (glykogensyntes i levern och förstärkning av matsmältningsprocesser); stärka processerna för urinering i njurarna och säkerställandet av urinering (sammandragning av blåsans muskler och avslappning av dess ringmuskel), etc. Det parasympatiska nervsystemet ger huvudsakligen startinfluenser : sammandragning av pupillen, bronkier, aktivering av aktivitet matsmältningskörtlar och så vidare.
Aktiviteten hos den parasympatiska uppdelningen av det autonoma nervsystemet är riktad mot strömmen reglering av det funktionella tillståndet, för att upprätthålla beständighet inre miljö homeostas . Den parasympatiska uppdelningen ger återställande av olika fysiologiska parametrar , drastiskt förändrats efter ansträngande muskelarbete påfyllning av förbrukade energiresurser. Medlare parasympatiska systemet acetylkolin, som minskar känsligheten hos adrenerga receptorer för verkan av adrenalin och noradrenalin, har en viss anti-stress effekt .
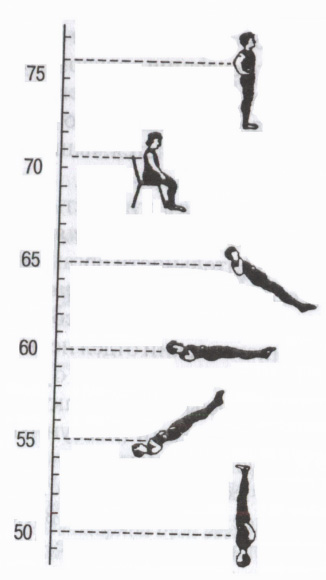
Ris. 5. Vegetativa reflexer
Effekt av kroppsposition på hjärtfrekvens (bpm). (Enligt: Mogendovich M.R., 1972)
Genom de autonoma sympatiska och parasympatiska vägarna utför det centrala nervsystemet några autonoma reflexer, som börjar med olika receptorer i den yttre och inre miljön: viscero-visceral (från inre organ till inre organ - till exempel andnings-hjärtreflexen); dermo-visceral (från huden - en förändring i aktiviteten hos inre organ under irritation aktiva punkter hud, såsom akupunktur, akupressur); från ögonglobens receptorer Ashners ögon-hjärtreflex (minskning av hjärtfrekvensen när du trycker på ögonglober parasympatisk effekt); motorisk-visceral-till exempel, ortostatiskt test(ökad hjärtfrekvens vid förflyttning från liggande till stående position - en sympatisk effekt) etc. (Fig. 5). De används för att bedöma kroppens funktionella tillstånd och särskilt tillståndet i det autonoma nervsystemet (bedöma påverkan av dess sympatiska eller parasympatiska avdelning).
limbiska systemet
Under limbiska systemet förstå ett antal kortikala och subkortikala strukturer vars funktioner är relaterade till organisation motiverande-emotionella reaktioner, minne och inlärningsprocesser .
De kortikala sektionerna av det limbiska systemet, som representerar dess högre sektion, är belägna på de nedre och inre ytorna av hjärnhalvorna (sektioner av frontala cortex, cingulate gyrus eller limbic cortex, hippocampus, etc.). TILL subkortikala strukturer Det limbiska systemet inkluderar hypotalamus, några kärnor i talamus, mellanhjärnan och retikulär formation. Mellan alla dessa avdelningar finns nära direkt- och återkopplingsförbindelser, som bildar den så kallade "limbiska ringen".
Det limbiska systemet är involverat i en mängd olika manifestationer av kroppens aktivitet i regleringen av ät- och drickbeteende, sömn-vakna cykeln, i processerna för att bilda ett minnesspår (avsättning och återhämtning från minnet), i utvecklingen av aggressiva defensiva reaktioner ger selektivt beteende. Hon skapar positiva och negativa känslor med alla motoriska, vegetativa och hormonella komponenter. Elektrisk stimulering av olika delar av det limbiska systemet genom implanterade elektroder (i djurförsök, i kliniken vid behandling av patienter) avslöjade närvaron av nöjescentra som genererar positiva känslor, och missnöje som skapar negativa känslor. Den isolerade irritationen av sådana punkter i den mänskliga hjärnans djupa strukturer orsakade uppkomsten av en känsla av "orsakslös glädje", "meningslös längtan", "oansvarig rädsla".
Funktioner av hjärnbarken
Hos högre däggdjur och människor är den ledande delen av CNS hjärnbarken.
Kortikala neuroner
Cortex är ett lager av grå substans som är 2-3 mm tjockt och innehåller i genomsnitt cirka 14 miljarder nervceller. Karakteristiskt för det är överflödet av internuronala förbindelser, vars tillväxt fortsätter upp till 18 år, och i vissa fall ännu längre.
Huvudtyperna av kortikala celler är pyramidala och stellate neuroner. stellate neuroner associerad med processerna för uppfattning av stimuli och föreningen av aktiviteterna hos olika pyramidala neuroner.
Pyramidala neuroner utföra den efferenta funktionen av cortex (främst genom pyramidkanalen) och viutricorcal interaktionsprocesser mellan avlägsna nervceller. De största pyramidcellerna, de gigantiska pyramiderna i Betz, finns i den främre centrala gyrusen (motorisk cortex).
Den funktionella enheten i cortex är vertikal kolumn sammankopplade neuroner. Vertikalt långsträckta stora pyramidceller med neuroner placerade ovanför och under dem bildar funktionella associationer av neuroner. Viktneuroner i den vertikala kolonnen svarar på samma afferenta stimulering (från samma receptor) med samma reaktion och bildar tillsammans efferenta svar av pyramidala neuroner. Vid behov kan vertikala kolonner kombineras till större formationer, vilket ger komplexa reaktioner.
Funktionell betydelse av olika kortikala fält
Enligt de strukturella egenskaperna och funktionella betydelsen av enskilda kortikala områden hela cortex är indelat i tre huvudgrupper av fält primär, sekundär och tertiär (Fig. 6).
Primära fält associerade med sinnesorganen och rörelseorganen i periferin. De bidrar förekomsten av förnimmelser . Dessa inkluderar till exempel smärtfältet och muskel-artikulär känslighet i den bakre centrala gyrusen av cortex, synfältet i den occipitalregionen, det auditiva fältet i den temporala regionen och det motoriska fältet i den främre centrala gyrusen. I de primära områdena finns högt specialiserade determinantceller eller detektorer, svarar selektivt endast på vissa stimuli. Till exempel i den visuella cortexen finns detektorneuroner som exciteras endast när ljuset slås på eller av, endast känsliga för dess bestämda intensitet, för specifika intervall av ljusexponering, för en viss våglängd, etc. Med förstörelsen av de primära fälten i cortex uppstår så kallad kortikal blindhet, kortikal dövhet etc.
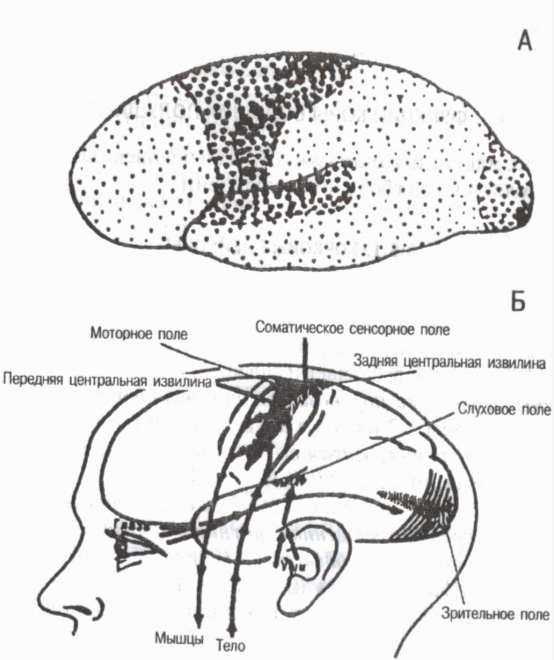
Ris. 6. Primära, sekundära och tertiära fält i hjärnbarken
På A: stora punkter primära fält, medelstora sekundära fält, små tertiära fält (enligt G. I. Polyakov 1964, A. R. Luria, 1971)
På B: primära (projektions)fält av hjärnbarken (enligt W. Penfield, L. Robert, 1964)
Sekundära fält ligger bredvid primären. Det händer i dem förståelse och igenkänning av ljud, ljus och andra signaler, komplexa former av generaliserad perception uppstår . När de sekundära fälten påverkas bevaras förmågan att se föremål, höra ljud, men personen känner inte igen dem, kommer inte ihåg innebörden.
Tertiära fält utvecklats nästan uteslutande hos människor. Dessa är associativa områden i cortex som tillhandahåller de högsta formerna av analys och syntes och bildar en persons målmedvetna beteendeaktivitet. Tertiära fält är belägna i den bakre halvan av cortex mellan parietal, occipital och temporal regioner och i den främre halvan i de främre delarna frontområden. Deras roll är särskilt viktig i organisationen av det samordnade arbetet för båda hemisfärerna . Tertiära fält mognar hos människor senare än andra kortikala fält och bryts ned tidigare än andra under åldrandet.
Funktionen hos de bakre tertiära fälten (främst de nedre parietalområdena i cortex) är mottagning, bearbetning och lagring av information. De bildar idén om kroppsschema och rymdschema ger rumslig orientering av rörelser. Främre tertiära fält (anterofrontala områden) utföra allmän reglering av komplexa former av mänskligt beteende, utforma avsikter och planer, program för godtyckliga rörelser och kontroll över deras genomförande . Utvecklingen av tertiära fält hos människor är associerad med talfunktion. tänker ( inre tal) är endast möjligt med den gemensamma aktiviteten av olika sensoriska system, föreningen av information från vilken sker i tertiära fält. Med medfödd underutveckling av tertiära fält kan en person inte behärska tal (uttalar bara meningslösa ljud) och till och med de enklaste motorikerna (kan inte klä sig, använda verktyg etc.).
Paraktivitet och dominans av hemisfärerna
Informationsbehandling genomförs som ett resultat parad aktivitet av båda hemisfärerna hjärna. Men som regel en av hemisfärerna är ledande dominant . De flesta med ledande höger hand(högerhänt) den vänstra hjärnhalvan är dominant, och den högra hjärnhalvan är underordnad (subdominant).
Vänster halvklot jämfört med högern har en finare neural struktur, en större rikedom av neuronala sammankopplingar, en mer koncentrerad representation av funktioner och Bättre förutsättningar blodtillförsel. I den vänstra dominanta hemisfären finns det motoriska centret för tal (Brocas centrum), vilket ger talaktivitet, och det sensoriska centrumet för tal, som förstår ord. Den vänstra hjärnhalvan är specialiserad på fin sensorimotorisk kontroll av handrörelser.
Människan har tre former funktionell asymmetri: motoriskt, sensoriskt och mentalt . Som regel har en person en ledande hand, fot, öga och öra. Problemet med funktionell asymmetri är dock ganska komplext. Till exempel kan en högerhänt person ha det dominerande vänstra ögat eller vänster öra, vars signaler är dominerande. Samtidigt, i varje halvklot, kan funktionerna hos inte bara den motsatta, utan också samma sida av kroppen representeras. Som ett resultat är det möjligt att ersätta en halvklot med en annan i händelse av skada, och skapar också strukturell grund för variabel hemisfärisk dominans vid motorisk kontroll.
Mental asymmetri visar sig i form av en viss hemisfäriska specialiseringar. För vänster hjärnhalva analytiska processer, sekventiell bearbetning av information, inklusive med hjälp av tal, abstrakt tänkande, bedömning av tidsmässiga förhållanden, förutsägelse av framtida händelser och framgångsrik lösning av verbal-logiska problem är karakteristiska. I högra hjärnhalvan information bearbetas holistiskt, syntetiskt (utan att dela upp i detaljer), med hänsyn till tidigare erfarenheter och utan deltagande av tal, råder objektivt tänkande. Dessa funktioner gör det möjligt att associera med den högra hjärnhalvan uppfattningen av rumsliga egenskaper och lösningen av visuell-spatiala uppgifter. Den högra hjärnhalvans funktioner är förknippade med det förflutna och den vänstra med framtiden.
Elektrisk aktivitet i hjärnbarken
Förändringar i det funktionella tillståndet hos cortex återspeglas i dess inspelning elektrisk aktivitet elektroencefalogram (EEG). Moderna elektroencefalografer förstärker hjärnans potentialer med 2-3 miljoner gånger och gör det möjligt att studera EEG från många punkter i cortex samtidigt, d.v.s. att studera systemprocesser. EEG-registrering gjorda i form av bläck på papper, såväl som i form komplett bild på hjärnans yta, d.v.s. kartor över hjärnan (kartläggningsmetod) på bildskärmen i moderna datorsystem (fig. 7).
Det finns vissa frekvensområden , ringde EEG rytmer (Fig. 8): i ett tillstånd av relativ vila registreras alfarytmen oftast (8-13 svängningar per 1 s); i ett tillstånd av aktiv uppmärksamhet - beta-rytm (14 oscillationer i 1s och över); när man somnar, vissa känslomässiga tillstånd betarytm (4-7 svängningar på 1s); med djup sömn, medvetslöshet, anestesi-delta rytm (1-3 fluktuationer på 1 s).
EEG speglar egenskaperna hos interaktionen av kortikala neuroner under mental och fysiskt arbete (Livanov M.N., 1972). Brist på koordination när man utför ovanligt eller hårt arbete leder till den sk EEG-desynkronisering snabb asynkron aktivitet. När en motorisk färdighet bildas uppstår fenomen i EEG EEG-synkronisering stärka sammankopplingen (synkron och i-fas) av den elektriska aktiviteten i olika områden av cortex involverade i kontrollen av rörelser. Under cykliskt arbete uppstår långsamma potentialer i takt med den utförda, imaginära eller kommande rörelsen "taggade rytmer"(Sologub E.B., 1973).
Utöver bakgrundsaktiviteten i EEG finns det separata händelserelaterade potentialer: framkallade potentialer uppstår som svar på yttre stimuli (auditiva, visuella, etc.); potentialer som återspeglar hjärnans processer under förberedelserna, genomförandet och slutförandet av individuella motoriska handlingar detta "våg av förväntan" eller en betingad negativ våg (Walter G., 1966), premotoriska, motoriska och slutgiltiga potentialer etc. Dessutom kan olika superlångsamma svängningar som varar från flera sekunder till tiotals minuter (särskilt de så kallade "omega-potentialerna", etc.), som återspeglar de biokemiska processerna för reglering av funktioner och mental aktivitet.

Ris. 7. Hjärnkartläggning
Flerkanalsinspelning av ett mänskligt elektroencefalogram (EEG) på en monitorskärm och reflektion av exciterade (ljuszoner) och hämmade (mörka zoner) områden i cortex.
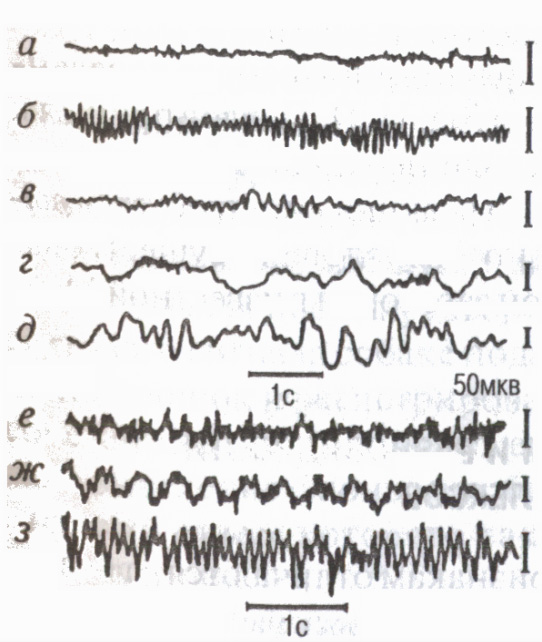
Ris. 8. EEG av de occipitala (a-e) och motoriska (f-h) områdena i den mänskliga hjärnbarken i olika stater och under muskelarbete
ett aktivt tillstånd,
öppna ögon (betarytm); b- vila, slutna ögon (alfarytm);
vid dåsighet (tesha-rytm);
d somna; e djup dröm"H
(delta rytm); f ovanligt eller
hårt arbete asynkront ofta
aktivitet (fenomen av desynkronisering);
g cykliskt arbete långsamma potentialer i takt med rörelser ("märkta rytmer" EEG); h framförande av den bemästrade rörelsen utseendet på alfarytmen.
Medicin och veterinär
pyramidala neuroner. stora celler, perikaryoner 10-130 mikron i storlek. Cellen har en apikal dendrit, som är riktad mot molekylskiktet; laterala processer - dendriter; ett långt axon går från basen - det kommer att vara en motorisk nervfiber (början på bildandet av pyramidkanalen).
Föreläsning om histologi №13
Centrala nervsystemet. Hjärnbarken. Cytoarchietektonik av skikten i hjärnbarken. neural sammansättning. Karakterisering av pyramidala neuroner. Modul strukturell och funktionell enhet av hjärnbarken. Lilla hjärnan. Strukturen och neuronala sammansättningen av cerebellar cortex.
Källa till utveckling av den grå och vita substansen i hjärnan
Utvecklas från manteln eller mantelzonen i neuralrörets grå substans
marginell slöja vit substans
Sammansatta komponenter grå substans i hjärnan (neocortex)
6 lager:
- Molekylär bildas huvudsakligen av processer av celler i de underliggande lagren, men det finns enstaka neuroner som kallas horisontella.
- Yttre granulär närvarande stellate neuroner och inte många pyramidala neuroner.
- Yttre pyramidal (pyramidal) det bredaste lagret som finns pyramidala neuroner.
- Inre granulär fin stellate neuroner, utför intrakortikala anslutningar (gå inte in i den vita substansen).
- Intern pyramidal (ganglion) representerad av stor pyramidal neuroner eller Beths-celler.
- Polymorfa cellformer är olika, cellerna är små, deras axoner sträcker sig in i den vita substansen.
stellate neuroner.Perikaryoner är runda eller triangulära till formen, 4-8 µm i diameter; axonet och dendriterna i dessa celler är involverade i intrakortikala förbindelser.
pyramidala neuroner.Stora celler, perikaryoner 10-130 mikron stora. Cellen har en apikal dendrit, som är riktad mot molekylskiktet; laterala processer dendriter; från basen kommer ett långt axon kommer att vara en motorisk nervfiber (början av bildandet av pyramidkanalen). Säkerheter kommer från axonet: de går till närliggande celler, eller reser sig upp och interagerar med de överliggande lagren.
generella egenskaper kortikala neuroner:
Alla neuroner i hjärnbarken är multipolära. Efter funktion, 1-4 lager: associativ (insert), 5-6 motor.
Strukturell och funktionell enhet i hjärnan: modul
Modul (i hjärnbarken upp till 3 miljoner) en vertikal kolumn med en diameter på 300 mikron, vinkelrät mot ytan av cortex och passerar genom hela tjockleken av cortex.
I mitten av modulen:
- En kortiko-kartisk nervfiber är en nervfiber i cortex; kommer antingen från pyramidala neuroner i samma hemisfär (associativ-interkalär), eller från den motsatta hemisfären (commissural).
- Två thalamuskortikala fibrer (affirent-sensoriska fibrer) som kommer från thalamus och går i 4 lager; då bildas en efferent nervfiber bildas av axoner Bästa celler.
Cytoarkitektoniksektioner av cortex, som skiljer sig från varandra när det gäller struktur och arrangemang av celler.
Myeloarkitektonikområden i cortex som skiljer sig från varandra i egenskaperna hos fibrernas placering.
fält olika områden i cortex som skiljer sig från varandra: cyto, myeloarkitektonik och funktionell betydelse.
Barktyper:
- Granulärt känsligt centrum, i vilket 2:a och 4:e lagret är väl utvecklade; de slutar i (afferenta) sensoriska nervfibrer som kommer från lukt-, hörsel- och synorganet.
- Agranulärt motorcentrum, där 3:e, 5:e och 6:e lagret utvecklas.
En pålitlig morfologisk motsvarighet till intelligens är antalet synapser av associativa neuroner (lager 1-4), normalt 10 tusen.
Lilla hjärnan
Källa till utveckling av cerebellums bakhjärna
Funktioner: koordinerar rörelse och balans
Lillhjärnan har ett stort antal veck, på beredningen i form av ett grenat träd.
Gyri är åtskilda av fåror. Varje gyrus innehåller: en smal platta av vit substans, helt täckt grå materia(bark). I djupet av den vita substansen i lillhjärnan ligger kluster av neuroner: kärnor i lillhjärnan.
Det finns tre lager i den grå substansen eller cerebellar cortex:
- External Molecular representeras av multipolära associativa (interkalära) neuroner. Det finns 2 typer av celler:
- Korgneuroner är belägna i den nedre tredjedelen av det molekylära lagret, celler oregelbunden form små storlekar. Cellens axon går in i en synaps med Purkenje-cellens perikareon, där den förgrenar sig för att bilda ett plexus i form av en korg. Dendriter i det molekylära lagret.
- Stjärnformade (stora och små) är lokaliserade till utsidan av det molekylära lagret (cortexytan).Stora stellate neuroner:axonet synapsar med Purkenje-cellernas perikaryon och deltar i bildandet av korgen. Dendriter i det molekylära lagret.små stjärnneuroner: Axonet synapsar med dendriterna i Purkenje-cellerna.
- Middle Ganglion - representeras av ett enda lager, kropparna av multipolära motorneuroner eller Purkinje-celler. Dessa är stora päronformade celler från perikaryon (kroppen) 2-3 dendriter sträcker sig in i det molekylära lagret. Axonet lämnar cellkroppen och passerar genom det granulära lagret in i den vita substansen, där den slutar vid de cerebellära kärnorna. Axoner av Purkinje-celler är de enda efferenta (motoriska) nervfibrerna som kommer ut från cerebellarbarken. Närmare cellkroppen avgår kollateraler från axonet, som går till ganglielagret och djupa delar av det molekylära lagret, där de går in i synaps med kroppar och dendriterangränsande Purkinje-celler.
- Inre granulär alla celler är polära och associativa innehåller två typer av celler:
1-Cells-Grains små celler, perikaryoner har en diameter på 4-6 mikron, dendriter: korta 3-4 stycken, riktade mot den vita substansen, som påminner om fågelben. Axonet stiger upp i det molekylära lagret, där det delar sig i en T-form.
2-Stor stjärnformad: med långa axoner och korta axoner Golgi-celler av den andra typen. Stora celler i kroppen är lokaliserade omedelbart under ganglielagret. Axonet går in i synaps med rosetter av mossiga fibrer innan de går in i synaps med granulcellsdendriter. Dendriterna stiger upp i det molekylära lagret.
Afferenta (sensoriska) nervfibrer:
- Bryofytfibrer Bryofytterminal nervfiber kallas uttag. Rosetter av den mossiga fibern synapsar med dendriterna av granulatceller och bildar cerebellära glomeruli.
- Klätter- eller lianliknande fibrer kommer från den vita substansen, passerar genom de granulära skikten och går in i en synaps antingen med Purkinjecellens perikaryon eller med dessa cellers dendriter.
Axoner av kornceller går in i synaps med dendriter:
- cellmolekylärt lager
- Cell Purkenye
- Golgi-celler av den andra typen
Det hämmande systemet i cerebellum inkluderar:
- Celler i det molekylära lagret
- Golgi-celler av den andra typen
- Excitatoriska synapser:
- Mossiga fibrer
- Lianoid fibrer
Samt andra verk som kan intressera dig |
|||
| 73026. | Mata in, formatera data och skriva formler | 126KB | |
| Syfte med arbetet: Använd kommandot Format Cell och formatera data i kolumn D med ett monetärt format utan decimaler. Använd knappen Format Painter och kopiera formatet för kolumn D till E. Använd knapparna på formateringsfältet för att ställa in kolumn F till ett procentformat med två decimaler. | |||
| 73027. | Bygga och redigera diagram i Excel | 160 kB | |
| Lär dig hur du bygger diagram med hjälp av guiden; Lär dig hur du redigerar diagram. Kontrollfrågor Vad är syftet med diagram Vilka typer av diagram vet du Hur man bygger ett diagram på separat blad Hur man ändrar diagramtypen Hur man tar bort ett diagram... | |||
| 73028. | Modellering av filsystem | 147,5 kB | |
| Användare ger filer symboliska namn, medan vissa OS-begränsningar beaktas. Katalogen innehåller en lista över filer som ingår i den och en överensstämmelse upprättas mellan filerna och deras egenskaper och attribut. | |||
| 73029. | Beteckningen av HBMs strukturfaslager, som ska kompensera för VNT, med metoderna för röntgendiffraktion och elektronmikroskopi | 1,5 MB | |
| Ange strukturfasstrukturen för HBM för att hämnas VNT för röntgendiffraktion och elektronmikroskopi. Följ upp förändringen av HBMs strukturella lager i termokemisk process. | |||
| 73033. | Anpassade datatyper på exemplet med MS SQL Server | 51 kB | |
| För att studera möjligheterna att definiera och använda anpassade datatyper, utlöser DML och DDL, integritetsbegränsningar i en postrelationell DBMS. Bemästra metoderna och teknikerna för att skapa anpassade datatyper på CLR.Net. | |||
| 73034. | Principen för märkning av armerade betonggolvplattor av civila byggnader | 47,1 kB | |
| Principen för märkning av armerade betonggolvplattor av civila byggnader I enlighet med GOST 2300978 är golvplattor märkta med ett antal alfanumeriska index. Den första gruppen innehåller plattans typbeteckning och dess konstruktionsmått, längd och bredd i decimeter. | |||
Nervsystem. Föreläsning 2
Bark stor hjärna (cortex cerebri) är ett kontinuerligt lager av grå substans på ytan av hjärnhalvorna med en tjocklek av 2-5 mm. Den ligger inte bara på vecken utan fortsätter in i alla fårorna, vilket gör att dess yta är relativt stor och uppgår till 2200 cm 2 hos en vuxen. Barkens massa är 581 g, dess volym är cirka 560 cm3. Samtidigt består skorpans huvudvolym av vit substans (450 cm 3, vikt - 470 g). Kroppsmassan hos kortikala neuroner är endast cirka 21 g (20 cm 3).
Storleken på kortikala nervceller varierar kraftigt från 8-9 mk upp till 150 mk. I den mänskliga hjärnbarken finns upp till 15 miljarder nervceller, varav 6 miljarder är små celler. De allra flesta kortikala neuroner är av två typer: pyramidala neuroner och stellateuroner. Dessa celler finns i cortex i vissa lager.
I den mänskliga hjärnan finns det flera fylogenetiskt olika typer bark:
Den gamla cortex - paleocortex (0,6%) - skiktningen är praktiskt taget frånvarande (1-2 lager), lokaliserad i regionen av lukttrianglarna, bildar ett täcke av den transparenta septumet och omger amygdala-kärnan.
Den gamla cortex - archicortex (2,2%) - har 2-3 lager, representeras av den grå substansen i dentate gyrus, den grå substansen i hippocampus och de cerebrala ränderna på corpus callosum.
Transitional cortex - mesokortex (1,6%) - området av den välvda gyrusen.
Den nya cortexen, neocortex (95,6%), är välstrukturerad och har 6 lager.
Hos högre ryggradsdjur finns det övervägande sex mer eller mindre tydligt definierade lager. Men vart och ett av dessa skikt, förutom det första, kan delas upp i två eller till och med tre underskikt.
Första lagret, den så kallade zon- eller molekylär platta, består huvudsakligen av plexus av apikala dendriter av pyramidala neuroner, vars cellkroppar är belägna i andra lager av cortex. Det finns väldigt få nervceller i det första lagret. Den presenterar horisontella celler, vars axoner och dendriter också är placerade horisontellt i samma lager.
Andra lagret, den så kallade externa granulära eller yttre granulär platta, inkluderar en massa små celler som tillhör små pyramidala, så kallade interkalära neuroner, och stellate neuroner, med små pyramidala celler som dominerar i det andra lagret.
Det tredje lagret är den yttre pyramidplattan innehåller medelstora pyramidceller
fjärde skiktet, den så kallade interna granulära eller inre granulär lamina, består huvudsakligen av små stjärnceller, men den innehåller även små och medelstora pyramider.
Femte lagret - den inre pyramidplattan innehåller gigantiska pyramidceller eller Betz-celler.
Sjätte lagret – multiform platta innehåller huvudsakligen pyramidformade celler av medelstora och en liten mängd små pyramid- och stellatceller.
De tre första lagren är de yngsta, de ger en koppling mellan olika delar av cortex. Det fjärde lagret är maximalt utvecklat i områden där afferent information kommer (känsliga centra, särskilt den postcentrala gyrusen). Det femte lagret uttrycks i de motoriska områdena i cortex: precentral gyrus, pericentral lobule, supramarginal gyrus.
Varje pyramidal neuron har en pyramidformad cell och många dendriter. Axonet av en pyramidal neuron kommer ut från en liten kulle vid basen av cellen. I små, så kallade interkalära eller mellanliggande pyramidala neuroner, slutar axoner, som förgrenar sig i horisontell eller vertikal riktning, omedelbart, utan att lämna cortex. Axoner av medelstora och stora pyramider avger många kollateraler i cortex, och huvudstammarna går in i den subkortikala vita substansen. Vissa av dem återvänder från den subkortikala substansen till cortex av en given hemisfär, eller, som passerar genom corpus callosum, slutar i cortex av den andra hemisfären. De tjänar till att förena olika delar av hjärnbarken. Därför kallas sådana pyramidala neuroner associativ. Andra axoner går till subkortikala formationer och vidare till olika delar av hjärnan och ryggmärgen. Dessa pyramider kallas utsprång. I samband med däggdjurens fylogenetiska utveckling ökar antalet pyramidceller kraftigt.
Bredden på hela cortex, antalet celler och bredden av varje lager av celler i frontal sektionen, såväl som den cellulära sammansättningen, det vill säga storleken, formen och arrangemanget av celler i varje sektion av cortex, varierar enormt. Dessa variationer upptar vissa, tydligt avgränsade territorier. I den stora hjärnan hos däggdjur finns det 11 väldefinierade stora kortikala fält.
På grundval av histologiska och fysiologiska studier delades varje sådant fält in i flera mer oberoende sektioner. En mer signifikant differentiering av cortex observeras hos högre däggdjur - apor. Det är särskilt bra hos människor.
Början av sådan forskning lades av Kiev-forskaren Vladimir Alekseevich Bets, som 1874 publicerade en artikel "Två centra i hjärnbarken", där han beskrev motorzonen i den precentrala gyrusen och den sensoriska zonen i den postcentrala.
1909 publicerade den tyske neurologen Korbinian Brodmann kartor över cytoarkitektoniska fält. hjärnbarken. Brodman var den första som skapade kartor över jordskorpan. Därefter beskrev O. Vogt och C. Vogt (1919-1920), med hänsyn till fiberstrukturen, 150 myeloarkitektoniska regioner i hjärnbarken. Vid Institute of the Brain of the USSR Academy of Medical Sciences skapade IN Filippov och SA Sarkisov kartor över hjärnbarken, inklusive 47 cytoarkitektoniska fält.
Data experimentella studier indikerar att förstörelsen eller avlägsnandet av vissa områden i hjärnbarken hos djur kränkte vissa funktioner. Dessa fakta bekräftas kliniska observationer för sjuka personer med tumörskador eller skador i vissa delar av hjärnbarken. Allt detta ledde till slutsatsen att det i hjärnbarken finns centra som reglerar utförandet av vissa funktioner. Morfologisk bekräftelse av data från fysiologi och klinik var läran om den heterogena strukturen av cortex i hjärnhalvorna i dess olika delar - cyto- och myeloarkitektonik i cortex. Det har fastställts att neuroner inte är belägna i cortex, utan är grupperade i ensembler.
Användningen av moderna mikroelektrodmetoder för att studera funktionerna hos kortikala neuroner har avsevärt utökat förståelsen för sensorisk informationsbehandling i neocortex och den strukturella organisationen av cortex. År 1957 upptäckte den amerikanske forskaren W. Mountcastle, som analyserade reaktionerna från celler i somatosensoriska (sensoriska motoriska) cortex hos en katt på stimuli av olika modaliteter, följande intressant fakta. När mikroelektroden var nedsänkt vinkelrätt mot ytan av den somatosensoriska cortex, reagerade alla celler som den mötte på samma sätt på en stimulans, till exempel på en lätt beröring av huden eller på rörelse i en led. Om elektroden var nedsänkt i en vinkel mot ytan av cortex, kom neuroner med olika sensorisk modalitet över längs dess väg, alternerande med en viss periodicitet.
Baserat på dessa experimentella fakta kom W. Mountcastle till slutsatsen att den somatosensoriska cortex är organiserad i elementära funktionella enheter - kolumner orienterad vinkelrätt mot ytan. Diametern på en sådan kolonn, cirka 500 µm, bestäms av den horisontella fördelningen av terminalerna på den afferenta talamokortikala fibern och den vertikala orienteringen av dendriterna. pyramidala celler. Enligt Mountcastle är kolonnen elementärt block av sensorimotorisk cortex, där lokal bearbetning av information från receptorer av samma modalitet utförs. Denna hypotes om den kolumnära organisationen av neocortex var allmänt accepterad och gav impulser till ytterligare forskning inom detta område i vårt land och utomlands.
Enligt moderna idéer Varje funktionell kolumn i den sensorimotoriska cortex består av flera morfologiska mikromoduler som kombinerar fem eller sex kapslade neuroner. Sammansättningen av en sådan modul inkluderar flera pyramidceller, vars apikala dendriter är så nära som möjligt och bildar en dendritisk bunt; inom denna stråle är möjliga elektrotoniska bindningar, som med all sannolikhet säkerställer en synkron drift av hela föreningen.
Intill en grupp av vertikalt orienterade pyramidceller finns stjärnceller med vilka de talamokortikala fibrerna som kommer till mikromodulen kommer i kontakt. En del av de hämmande stjärncellerna har långa axoner som sträcker sig horisontellt. Axoner av pyramidceller bildar återkommande kollateraler, vilket kan ge både underlättande influenser inom mikromodulen och hämmande interaktioner mellan mikromoduler genom att kontakta hämmande interneuroner. Flera strukturella mikromoduler, förenade genom horisontell förgrening av terminalerna av specifika thalamokortikala afferenter, axoner av terminalerna och processer av stellatceller, bildar en kolumn (eller makromodul), vars diameter når 500-1000 µm. Varje kolumn kännetecknas av funktionell enhet, vilket manifesteras i det faktum att nervcellerna i kolonnen svarar på en stimulans av en modalitet.
Därefter bekräftades principen om kolumnär organisation i studien av andra projektionszoner bark.
I.P. Pavlov betraktade hjärnbarken som en kontinuerlig uppfattningsyta, som en uppsättning kortikala ändar av analysatorer. Han visade att den kortikala änden av analysatorerna inte är en strikt definierad zon. I hjärnbarken urskiljs en kärna och spridda element. Kärnan är platsen för koncentrationen av nervceller i cortex, som utgör en exakt projektion av alla element i en viss perifer receptor, där den högsta analysen, syntesen och integrationen av funktioner äger rum. Spridda element kan lokaliseras både längs kärnans periferi och på ett betydande avstånd från den. De är enklare analys och syntes. Närvaron av spridda element under förstörelsen av kärnan gör det delvis möjligt att kompensera för den försämrade funktionen. Områden upptagna av spridda element olika analysatorer, kan överlappa varandra, överlappa varandra. Således kan hjärnbarken schematiskt föreställas som en uppsättning kärnor av olika analysatorer, mellan vilka det finns spridda element relaterade till olika (intilliggande) analysatorer. Ovanstående låter oss prata om dynamisk lokalisering av funktioner i hjärnbarken (I.P. Pavlov).
Låt oss överväga positionen för några kortikala ändar av olika analysatorer (kärnor) i förhållande till veck och lober i hjärnhalvorna hos människor i enlighet med cytoarkitektoniska kartor.
1. I cortex av den postcentrala gyrusen och den övre parietallobulien finns nervceller som utgör kärnan i den kortikala analysatorn hos den allmänna känslighet(temperatur, smärta, taktil) och proprioceptiva. Ledande sensoriska vägar som följer till hjärnbarken skär varandra antingen på nivån för olika segment av ryggmärgen (vägar av smärta, temperaturkänslighet, beröring och tryck), eller på nivån av medulla oblongata (vägar för proprioceptiv känslighet i den kortikala riktningen) ). Som ett resultat är den postcentrala gyri av var och en av hemisfärerna anslutna till den motsatta halvan av kroppen. I den postcentrala gyrusen projiceras alla receptorfält i olika delar av människokroppen på ett sådant sätt att de kortikala ändarna av känslighetsanalysatorn i de nedre delarna av kroppen är placerade högst och nedre extremiteterna, och den lägsta (närmare den laterala fåran) är receptorfälten i de övre delarna av kroppen och huvudet, de övre extremiteterna.
2. Kärna motor analysatorn är huvudsakligen placerad i det så kallade motoriska området av cortex, vilket inkluderar den precentrala gyrusen och den paracentrala lobulen på mediala ytan hemisfär. I det 5:e lagret av cortex av precentral gyrus ligger pyramidala neuroner (Betz-celler), som I.P. Pavlov kallade intercalary och noterade att dessa celler, med sina processer, är kopplade till de subkortikala kärnorna, motorcellerna i kärnorna i kraniet och spinal nerver. Dessutom, i de övre delarna av den precentrala gyrusen och i den paracentrala lobulen finns det celler, varifrån impulserna riktas till musklerna i de nedre delarna av bålen och nedre extremiteter. I den nedre delen av den precentrala gyrusen finns de motoriska centra som reglerar aktiviteten i ansiktets muskler.
Således projiceras alla delar av människokroppen upp och ner i den precentrala gyrusen. På grund av det faktum att pyramidvägarna, som kommer från jättelika pyramidceller, korsar antingen i nivå med hjärnstammen (kortikala-kärnfibrer), vid gränsen till ryggmärgen eller i segment av ryggmärgen (kortikal-ryggradsfibrer). tarmkanalen), är de motoriska områdena på var och en av halvkloten anslutna till skelettmusklerna på den motsatta sidan av kroppen. Om musklerna i extremiteterna är isolerade till en av hemisfärerna, är musklerna i bålen, struphuvudet och svalget anslutna till de motoriska områdena i båda hemisfärerna.
Motor I.P. Pavlov kallade också regionen i hjärnbarken för receptorområdet, eftersom här analysen av proprioceptiva (kinetiska stimuli uppfattas av receptorer inbäddade i skelettmuskler, senor, fascia och ledkapslar).
3. Analysatorns kärna som tillhandahåller funktionen kombinerad rotation av huvudet och ögonen in motsatta sidan , är i bakavdelningar mellersta frontal gyrus, i den så kallade premotoriska zonen.
4. I området för den nedre parietala lobulen, i den supramarginala gyrusen, finns kärnan av motoranalysatorn, vars funktionella betydelse är att implementera syntes Allt målmedvetna, professionella, arbetar- och idrottsrörelser. Denna kärna är asymmetrisk. För högerhänta är det i vänster och för vänsterhänta- endast i höger hjärnhalva. Förmågan att koordinera dessa komplexa målmedvetna rörelser förvärvas av en individ under sitt liv som ett resultat av praktisk aktivitet och ackumulering av erfarenhet. Implementeringen av målmedvetna rörelser uppstår på grund av bildandet av tillfälliga förbindelser mellan celler belägna i den precentrala och supramarginala gyrusen. Fältets nederlag orsakar inte förlamning, utan leder bara till förlust av förmågan att producera komplexa koordinerade målmedvetna rörelser - apraxi (praxis - praktik).
5. I cortex av den övre parietallobuli är kärnan i hudanalysatorn av en av de speciella typerna av känslighet, som har funktionen känna igen föremål genom beröring,- streognosi. Den kortikala änden av denna analysator är belägen i den högra hjärnhalvan. Nederlaget för ytskikten av cortex i detta avsnitt åtföljs av förlusten av funktionen att känna igen föremål genom beröring, även om andra typer av allmän känslighet bevaras.
6. I djupet av lateral sulcus, på ytan av den mellersta delen av den övre temporala gyrusen som är vänd mot holmen (där den transversella temporala gyrusen, eller Geschls gyrus) är synlig, finns en kärna auditiv analysator. TILL nervceller, som utgör kärnan hörselanalysator var och en av hemisfärerna, vägarna från receptorerna för både vänster och höger sida. I detta avseende orsakar en ensidig lesion av denna kärna inte en fullständig förlust av förmågan att uppfatta ljud. Bilateral lesion åtföljs av kortikal dövhet.
7. Kärna visuell analysatorn finns i occipitala loben cerebrala hemisfärer i omkretsen av sporrspåret. Kärna visuell analysator av höger hjärnhalva är förbunden med vägar med den laterala halvan av näthinnan i det högra ögat och den mediala halvan av näthinnan i det vänstra ögat. I cortex av den vänstra hemisfärens occipitallob projiceras receptorerna från den laterala halvan av näthinnan i det vänstra ögat och den mediala halvan av näthinnan i det högra ögat. När det gäller hörselanalysatorns kärna, leder endast bilateral skada på kärnorna i den visuella analysatorn till fullständig kortikal blindhet. Fältets nederlag, som är något högre än kärnan i den visuella analysatorn, åtföljs av en förlust av visuellt minne, men ingen synförlust noteras. Ännu högre är en del av cortex, vars nederlag åtföljs av en förlust av förmågan att navigera i en obekant miljö.
8. På den nedre ytan av tinningloben i hjärnhalvan, i området av kroken och delvis i området av hippocampus, finns en kärna lukt- analysator. Dessa områden, från fylogenessynpunkt, tillhör de äldsta delarna av hjärnbarken. Lukt- och smaksinnet är nära besläktade, vilket förklaras av den nära platsen för kärnorna i lukt- och smakanalysatorerna. Det noterades också (V.M. Bekhterev) att smakuppfattningen störs när cortex i de lägsta delarna av den postcentrala gyrusen är skadad. Kärnorna i smak- och luktanalysatorerna i båda hemisfärerna är associerade med receptorer på både vänster och höger sida av kroppen.
De beskrivna kortikala ändarna av vissa analysatorer finns inte i hjärnbarken. bara människor, men också djur. De är specialiserade på perception, analys och syntes av signaler som kommer från den externa och interna miljön, som enligt IP Pavlovs definition utgör verklighetens första signalsystem.
Andra signalsystemet finns endast hos människor och beror på talets utveckling. Tal, och medvetandet tillsammans med det, är fylogenetiskt de yngsta funktionerna i hjärnan. I detta avseende är de kortikala ändarna av analysatorerna minst lokaliserade. Tal och mentala funktioner utförs med deltagande av hela cortex. Vissa områden kan dock urskiljas i hjärnbarken, som kännetecknas av strikt definierade talfunktioner. Ja, analysatorer motoriskt tal(oralt och skriftligt) är belägna bredvid det motoriska området av cortex, närmare bestämt i de delar av cortex av frontalloben som gränsar till den precentrala gyrusen.
Analysatorer visuella och hörseluppfattning talsignaler finns bredvid syn- och hörselanalysatorerna. Det bör påpekas att talanalysatorerna hos högerhänta personer är lokaliserade i vänster hjärnhalva och hos vänsterhänta - främst i höger.
Tänk på positionen i hjärnbarken hos några av talanalysatorerna.
1. Motoranalysatorn för skriftligt tal placeras i den bakre delen av den mellersta frontala gyrusen. Aktiviteten hos denna analysator är nära förbunden med analysatorn av målmedvetna rörelser, som säkerställer bildandet av de inlärda rörelserna i handen och ögonen som är nödvändiga för att skriva. Vid skada förloras förmågan att utföra fina rörelser, vanligtvis utförda under visuell kontroll och nödvändiga för inskription av bokstäver, ord och andra tecken (agraphia).
2. Kärnan i motoranalysatorn talartikulation(talmotoranalysator) är placerad i de bakre sektionerna av den nedre frontala gyrusen (Brocas mitt). Skador på cortex i detta område leder till motorisk afasi, det vill säga förlust av förmågan att uttala ord (afasi). Denna afasi är inte förknippad med en förlust av förmågan att dra ihop de muskler som är involverade i talproduktion. Dessutom, med nederlaget för detta fält, förloras inte förmågan att uttala ljud eller sjunga.
3. In centrala avdelningar den nedre frontala gyrusen är belägen kärnan i talanalysatorn förknippad med sång. Hans nederlag åtföljs av vokalamusia - oförmågan att komponera och reproducera musikaliska fraser och agrammatism, när förmågan att komponera meningsfulla meningar från enskilda ord går förlorad. Sådana människors tal består av en uppsättning ord som inte är relaterade i semantisk betydelse.
4. Kärna auditiv talanalysator nära besläktad med
kortikala mitten av den auditiva analysatorn och är belägen i de bakre sektionerna
överlägsen temporal gyrus, på dess yta vänd bort
lateral sulcus av hjärnhalvan. Skador på kärnan försämrar inte hörseluppfattningen av ljud i allmänhet, men förmågan att förstå ord och tal går förlorad (verbal dövhet eller sensorisk afasi). Funktionen hos denna kärna är att en person inte bara hör och förstår en annan persons tal, utan också kontrollerar sitt eget.
5. I den mellersta tredjedelen av den överlägsna temporala gyrusen finns kärnan i den kortikala analysatorn, vars nederlag åtföljs av uppkomsten av musikalisk dövhet, när musikaliska fraser uppfattas som en meningslös uppsättning olika ljud. Denna kortikala ände av den auditiva analysatorn tillhör centra för det andra signalsystemet, som uppfattar den verbala beteckningen av objekt, handlingar, fenomen, d.v.s. ta emot signaler.
6. I direkt anslutning till kärnan av den visuella analysatorn är kärnan visuell analysator av skriftligt tal(fält 39), belägen i den vinkelformade gyrusen av den nedre parietala lobulen. Nederlaget för denna kärna leder till förlust av förmågan att uppfatta den skrivna texten, att läsa (dyslexi).
En persons frontala cortex är förknippad med en social princip. Dess neuroner bildar tvåvägsförbindelser med alla andra delar av cortex. Överträdelser uppstår när de skadas socialt beteende, elakhet, illojalitet, minskad målmedvetenhet, apati. Efter lobotomi - en fullständig brist på initiativ och social aktivitet.
Asymmetrin av funktionerna i höger och vänster hemisfär är mycket uttalad. Den vänstra hjärnhalvan ger logik, abstrakt tänkande, dominerar i förhållande till talfunktioner (läsa, skriva, räkna), komplexa frivilliga rörelser. Höger hjärnhalva- emotionalitet, figurativt tänkande, icke-talfunktioner för igenkänning av komplexa visuella och auditiva bilder, taktila perceptioner, perception av rymd, form, riktning, intuitivt tänkande.
Liknande artiklar
-
När en man är emot ett barn, hur blir man gravid utan hans vetskap?
Ibland kan man bli gravid av oaktsamhet. För att förhindra att detta händer är det viktigt att veta hur du kan bli gravid med ett barn av misstag och vilka medel du kan använda för att undvika oönskad graviditet. Även i den här artikeln kan du hitta information om ...
-
Vilka stenar och amuletter är lämpliga för Oxen enligt horoskopet och födelsedatumet Elefanttalisman för Oxen
April-maj Oxen (21 april - 20 maj) är mätt, inte kinkig och kolossalt produktiv! Deras avundsvärda envishet kan få andra att ta tag i, men de vet exakt vad de gör och varför de behöver det. Bland de positiva...
-
Restriktioner för åtkomst till data i roller 1c
Alla användarrättighetsinställningar som vi kommer att göra inom ramen för denna artikel finns i avsnitt 1C 8.3 "Administration" - "Användar- och rättighetersinställningar". Denna algoritm är liknande i de flesta konfigurationer på ...
-
1c startar en tunn klient istället för en tjock
Plattformar: 1C: Enterprise 8.3, 1C: Enterprise 8.2, 1C: Enterprise 8.1 Konfigurationer: Alla konfigurationer2012-11-16 21362 De lanseras genom att specificera speciella ...
-
Bevis på kända sätt att stjäla el Hur man tar reda på vem som stjäl el
Höjningen av energitaxor är ett av de slående dragen i den fördjupade ekonomiska krisen. I detta sammanhang är stöld av el och de problem som är förknippade med dess upptäckt av största vikt. Sätt att upptäcka stöld ...
-
Funktioner för montering av uttag och strömbrytare på olika ytor
Hälsningar till alla läsare av vår blogg Idag, kära läsare, vill jag lyfta fram ämnet om hur man installerar uttag. Denna procedur efterfrågas mycket ofta när man byter ut ett gammalt uttag med ett nytt i händelse av ett haveri, när ...