Nervimpuls. nervimpulser
Neuroner kommunicerar med varandra med hjälp av "neurala meddelanden". Dessa "meddelanden" är som elektrisk ström som går genom ledningar. Ibland, när de överförs från en neuron till en annan, omvandlas dessa impulser till kemiska meddelanden.
nervimpulser
Information överförs mellan neuroner som en elektrisk ström i ledningar. Dessa meddelanden är kodade: de är en sekvens av exakt samma impulser. Själva koden ligger i deras frekvens, det vill säga i antalet pulser per sekund. Impulserna överförs från cell till cell, från dendriten där de har sitt ursprung till axonet genom vilket de passerar. Men det finns också en skillnad från elektriska nätverk - impulser överförs inte med hjälp av elektroner *, utan med hjälp av mer komplexa partiklar - joner.
Mediciner som påverkar impulshastigheten
Det är många kemikalier som kan förändra egenskaperna hos överföringen av nervimpulser. Som regel agerar de på synaptisk nivå. Bedövningsmedel och lugnande medel bromsar, och ibland till och med undertrycker, överföringen av impulser. Och antidepressiva och stimulantia som koffein bidrar tvärtom till deras bättre överföring.
Med stor fart
Nervimpulser måste färdas snabbt genom kroppen. Gliaceller som omger dem hjälper till att påskynda deras passage till neuroner. De bildar manteln av nervfibern, som kallas myelin. Som ett resultat går impulserna i hisnande hastighet - mer än 400 km/h.
kemiska bindningar
Meddelanden som överförs från neuron till neuron måste omvandlas från elektrisk till kemisk form. Detta beror på det faktum att nervceller, trots sitt stora antal, aldrig vidrör varandra. Men elektriska impulser kan inte överföras om det inte finns fysisk kontakt. Därför använder neuroner ett speciellt system som kallas synapser för att kommunicera med varandra. På dessa ställen är neuronerna åtskilda av ett smalt utrymme i den synaptiska klyftan. När en elektrisk impuls kommer till den första neuronen frigör den kemiska molekyler som kallas neurotransmittorer från synapsen. Dessa ämnen, som produceras av neuroner, rör sig genom den synaptiska klyftan och går in i receptorerna hos en annan neuron speciellt utformad för dem. Resultatet är ytterligare en elektrisk impuls.
En impuls färdas mellan neuroner på mindre än en tusendels sekund.
Särskiljande neurotransmittorer
Hjärnan producerar ett femtiotal signalsubstanser, som kan delas in i två grupper. Den första består av de som initierar förekomsten av en nervimpuls - de kallas excitatoriska. Andra, tvärtom, saktar ner dess förekomst - dessa är hämmande neurotransmittorer. Det bör noteras att i de flesta fall släpper neuronen endast en typ av signalsubstanser. Och beroende på om den är excitatorisk eller hämmande har neuronen olika effekt på närliggande nervceller.
konstgjord stimulering
En enskild neuron eller en grupp av neuroner kan stimuleras artificiellt med hjälp av elektroder som sätts in i dem, som riktar elektriska impulser till exakt markerade områden i hjärnan. Denna metod används ibland inom medicinen, särskilt för behandling av patienter som lider av Parkinsons sjukdom.Denna sjukdom, som visar sig i hög ålder, åtföljs av darrningar i armar och ben. Denna skakning kan stoppas genom konstant stimulering av ett specifikt område av hjärnan.
Neuron - mikrodator
Var och en av neuronerna kan ta emot hundratals meddelanden per sekund. Och för att inte bli överbelastad med information måste han kunna bedöma graden av dess betydelse och göra en preliminär analys av den. Denna beräkningsaktivitet äger rum inuti cellen. Excitatoriska impulser läggs till och hämmande impulser subtraheras. Och för att neuronen ska generera sin egen impuls, är det nödvändigt att summan av de föregående är större än visst värde. Om tillägget av excitatoriska och hämmande impulser inte överskrider denna gräns, kommer neuronen att vara "tyst".
informationsvägar
I alla dessa förvecklingar av neuroner finns det vackert markerade stigar. Liknande idéer, liknande minnen passerar, alltid avfyrar samma nervceller och synapser. Det är fortfarande okänt hur dessa kretsliknande elektroniska kommunikationskretsar skapas och underhålls, men det är uppenbart att de finns och att ju starkare de är desto effektivare är de. Ofta använda synapser fungerar snabbare. Detta förklarar varför vi snabbt kommer ihåg saker som vi har sett eller upprepat flera gånger. Dessa bindningar varar dock inte för evigt. Vissa av dem kan försvinna om de inte används tillräckligt, och nya kommer att dyka upp i deras ställe. Vid behov kan neuroner alltid skapa nya kopplingar.
Små gröna prickar på bilden - hormoner inuti blodkärl
Kemisk dopning
När de säger att en idrottare använde hormonell dopning betyder det att han tagit hormoner antingen i form av tabletter eller genom att injicera dem direkt i blodet. Hormoner är antingen naturliga eller konstgjorda. De vanligaste är tillväxthormoner och steroider, på grund av vilka musklerna blir större och starkare, samt erytropoietin, ett hormon som påskyndar förlossningen näringsämnen till musklerna.
Hjärnan är kapabel att utföra miljontals operationer på en bråkdel av en sekund.
Hormoner verkar på hjärnan
För utbyte av information av hjärnan används också ett annat verktyg - hormoner. Dessa kemiska föreningar produceras delvis av hjärnan själv i en grupp nervceller som finns i hypotalamus. Dessa hormoner styr produktionen av andra som produceras på andra ställen i kroppen i de endokrina körtlarna. De fungerar annorlunda än signalsubstanser, som är fixerade direkt på nervceller och transporteras i blodet till avlägsna organ i kroppen från hjärnan, såsom bröst, äggstockar, manliga testiklar och njurar. Genom att fixera på sina receptorer orsakar hormoner olika fysiologiska reaktioner. De främjar till exempel tillväxten av ben och muskler, kontrollerar känslan av hunger och törst och påverkar naturligtvis sexuell aktivitet.
NERVIMPULS- en excitationsvåg, som sprider sig längs nervfibern och tjänar till att överföra information från den perifera. receptor (känsliga) ändar till nervcentra, inuti centrum. nervsystemet och från det till den verkställande apparaten - musklerna och körtlarna. N:s gång och. åtföljs av transient elektrisk. processer, to-rye är det möjligt att registrera både extracellulära och intracellulära elektroder.
Generering, överföring och bearbetning N. och. Utför nervsystem. Main ett strukturellt element i nervsystemet hos högre organismer är en nervcell, eller en neuron, bestående av en cellkropp och många. processer - dendriter (Fig. 1). En av processerna i icke-rifärisk. neuroner har en stor längd - detta är en nervfiber eller axon, vars längd är ~ 1 m och tjockleken är från 0,5 till 30 mikron. Det finns två klasser nervfibrer: köttig (myeliniserad) och icke-köttlös. De massaliknande fibrerna har ett myelinhölje bildat av special. ett membran, kanter som isolering är lindade på ett axon. Längden på sektioner av en kontinuerlig myelinskida är från 200 mikron till 1 mm, de avbryts av den så kallade. avlyssningar av Ranvier med en bredd av 1 μm. Myelinskidan spelar rollen som isolering; nervfibern i dessa områden är passiv, endast membranet i Ranviers noder är elektriskt aktivt. Meleless fibrer har inte isolerat. tomter; deras struktur är homogen längs hela längden, och membranet har en elektrisk. aktivitet över hela ytan.
Nervfibrer slutar på andra nervcellers kroppar eller dendriter, men separeras från dem av en mellanliggande
en kuslig bredd på ~10 nm. Detta kontaktområde mellan två celler kallas. synaps. Axonmembranet som kommer in i synapsen kallas. presynaptisk, och motsvarande dendritiska eller muskelmembran är postsynaptisk (se fig. cellstrukturer).
I normala förhållanden en serie av N. och. som ständigt löper längs nervfibern, uppstår på dendriterna eller cellkroppen och sprider sig längs axonet i riktning från cellkroppen (axonen kan leda N. och. i båda riktningarna). Frekvensen av dessa periodiska utsläpp bär information om styrkan av irritationen som orsakade dem; till exempel när måttlig aktivitet frekvens ~ 50-100 pulser/s. Det finns celler, to-rye urladdas med en frekvens på ~ 1500 impulser/s.
Distributionshastigheten för N. och. u beror på typen av nervfiber och dess diameter d, u~ d 1/2. I tunna fibrer i det mänskliga nervsystemet, u ~ 1 m/s, och i tjocka fibrer, u ~ 100-120 m/s.
Varje N. och. uppstår som ett resultat av irritation av kroppen av en nervcell eller nervfiber. N. och. har alltid samma egenskaper (form och hastighet) oavsett styrka av irritation, d.v.s. med subtröskelstimulering av N. och. förekommer inte alls, men med suprathreshold - har full amplitud.
Efter excitation uppstår en refraktär period, under vilken nervfiberns excitabilitet reduceras. Särskilja abs. den refraktära perioden, då fibern inte kan exciteras av några stimuli, och refererar. refraktärperioden, när excitation är möjlig, men dess tröskel är över det normala. Magmuskler. refraktärperioden begränsar sändningsfrekvensen för N. uppifrån och. Nervfibern har egenskapen av ackommodation, det vill säga den vänjer sig vid ständigt verkande irritation, vilket uttrycks i en gradvis ökning av tröskeln för excitabilitet. Detta leder till en minskning av N:s frekvens och. och till och med deras fullständiga försvinnande. Om stimuleringsstyrkan ökar långsamt, kan det hända att excitation inte inträffar även efter att tröskeln har nåtts.
Figur 1. Diagram över strukturen av en nervcell.
Längs N:s nervfiber och. fortplantar sig som en elektrisk våg. potential. I synapsen sker en förändring i fortplantningsmekanismen. När N. och. når det presynaptiska ändelser, i synaptisk. gapet tilldelas aktiv kemisk. substans - m e d i a t o r. Mediatorn diffunderar genom synaptiken. gap och ändrar permeabiliteten för postsynaptisk. membranet, som ett resultat av vilket en potential uppstår på det, återigen genererar en fortplantningsimpuls. Så här fungerar kemoterapi. synaps. Det finns också en elektrisk synaps när nästa. neuronen är elektriskt exciterad.
N:s upphetsning och. Phys. idéer om utseendet på elektriska. potentialer i celler är baserade på den sk. membranteori. Cellmembran separerar elektrolytlösningar av olika koncentrationer och har iso-birat. permeabilitet för vissa joner. Således är axonmembranet ett tunt lager av lipider och proteiner med en tjocklek på ~7 nm. Hennes elektriska motstånd i vila ~ 0,1 ohm. m 2, och kapaciteten är ~ 10 mf / m 2. Inuti axonet är koncentrationen av K + joner hög och koncentrationen av Na + och Cl - joner är låg, och i miljö- vice versa.
I vila är axonmembranet permeabelt för K+-joner. På grund av skillnaden i koncentrationer C 0 K i ext. och C i ext. lösningar etableras en kaliummembranpotential på membranet
![]()
Var T- magmuskler. pace-pa, e- elektron. På axonmembranet observeras verkligen en vilopotential på ~ -60 mV, motsvarande den indikerade f-le.
Joner Na + och Cl - penetrerar membranet. För att upprätthålla den nödvändiga icke-jämviktsfördelningen av joner använder cellen ett aktivt transportsystem, vilket kräver cellulär energi. Därför är vilotillståndet för nervfibern inte termodynamiskt jämvikt. Den är stationär på grund av verkan av jonpumpar, och membranpotentialen i öppna kretsförhållanden bestäms från lika med noll av den totala elektriska. nuvarande.
Processen med nervös excitation utvecklas enligt följande (se också Biofysik Om en svag strömpuls leds genom axonet, vilket leder till depolarisering av membranet, sedan efter avlägsnande av det yttre. exponeringspotentialen återgår monotont till den ursprungliga nivån. Under dessa förhållanden beter sig axonet som en passiv elektrisk krets. krets bestående av en kondensator och en DC. motstånd.
Ris. 2. Utveckling av aktionspotentialen i nervsystemetlokne: A- undertröskel ( 1
) och övertröskel (2) irritation; b-membransvar; med irritation över tröskeln uppträder full svetthandlingscykel; Vär jonströmmen som flyter igenom membran när det exciteras; G- uppskattning jonström i en enkel analytisk modell.
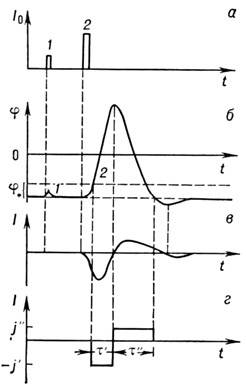
Om den aktuella pulsen överskrider ett visst tröskelvärde, fortsätter potentialen att förändras även efter att störningen stängts av; potentialen blir positiv och återgår först därefter till vilonivån, och först hoppar den till och med över lite (hyperpolariseringsregionen, fig. 2). Membranets respons beror inte på störningen; denna impuls kallas agerande potential. Samtidigt flyter en jonström genom membranet, riktad först inåt och sedan utåt (Fig. 2, V).
Fenomenologiska tolkning av mekanismen för förekomst av N. och. gavs av A. L. Hodg-kin och A. F. Huxley 1952. Den totala jonströmmen består av tre komponenter: kalium, natrium och läckström. När membranpotentialen skiftas med tröskelvärdet j* (~ 20mV) blir membranet permeabelt för Na+-joner. Na+-joner rusar in i fibern och förskjuter membranpotentialen tills den når natriumjämviktspotentialen:
![]()
komponent ~ 60 mV. Därför når aktionspotentialens fulla amplitud ~ 120 mV. När max. potential i membranet börjar utveckla kalium (och samtidigt minska natrium) ledningsförmåga. Som ett resultat ersätts natriumströmmen med en kaliumström riktad utåt. Denna ström motsvarar en minskning av aktionspotentialen.
Det empiriska ur-tion för beskrivning av natrium- och kaliumströmmar. Membranpotentialens beteende under rumsligt homogen excitation av fibern bestäms av ekvationen:
Var MED- membrankapacitans, jag- jonström, bestående av kalium, natrium och läckström. Dessa strömmar bestäms av posten. emf jK, j Na och j l och konduktiviteter g K , g Na och gl:
värdet gl anses vara konstant, konduktivitet g Na och g K beskrivs med hjälp av parametrar m, h Och P:
g Nej, g K - konstanter; alternativ t, h Och P uppfylla de linjära ekvationerna
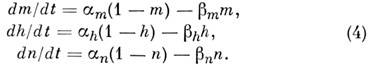
Koefficientberoende. a och b på membranpotentialen j (fig. 3) väljs från tillståndet för den bästa matchningen
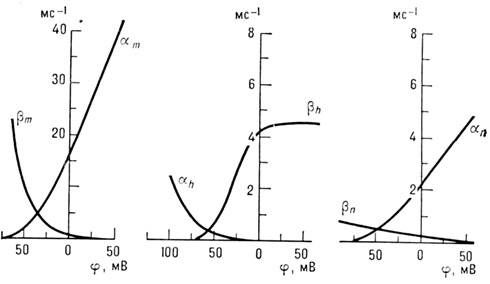
Ris. 3. Beroende av koefficienter a Och b från membrankapacitet.
beräknade och uppmätta kurvor jag(t). Valet av parametrar beror på samma överväganden. Beroende av stationära värden t, h Och P på membranpotentialen visas i fig. 4. Det finns modeller med ett stort antal parametrar. Således är nervfibermembranet en icke-linjär jonledare, vars egenskaper väsentligt beror på det elektriska. fält. Mekanismen för generering av excitation är dåligt förstådd. Hodgkin-Huxley Urn ger bara en framgångsrik empiri. beskrivning av fenomenet, för vilket det inte finns någon specifik fysisk. modeller. Det är därför viktig uppgiftär studiet av mekanismerna för flödet av elektriska. ström genom membran, särskilt genom kontrollerad elektrisk. fältjonkanaler.
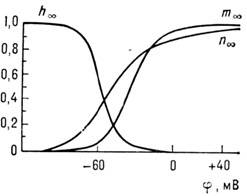
Ris. 4. Beroende av stationära värden t, h Och P
från membranpotential.
N:s fördelning och. N. och. kan fortplanta sig längs fibern utan dämpning och med stolpe. fart. Detta beror på att den energi som behövs för signalöverföring inte kommer från ett enda centrum, utan dras på plats, vid varje punkt av fibern. I enlighet med de två typerna av fibrer finns det två sätt för N:s överföring och
Vid icke-myelinisering fibermembranpotentialfördelning j( x, t) bestäms av ekvationen:
Var MEDär kapacitansen för membranet per enhet fiberlängd, Rär summan av longitudinella (intracellulära och extracellulära) motstånd per enhet fiberlängd, jag- jonström som flyter genom membranet hos en fiber av enhetslängd. Elektrisk nuvarande jagär en funktion av potentialen j, som beror på tiden t och koordinater X. Detta beroende bestäms av ekvationerna (2) - (4).
Typ av funktionalitet jag specifikt för en biologiskt exciterbar miljö. Emellertid, ekvation (5), bortsett från formen jag, har mer allmän karaktär och beskriver många fysiska fenomen, t.ex. förbränningsprocess. Därför N:s överlåtelse och. liknas vid bränning av en pulversnöre. Om i en rinnande låga antändningsprocessen utförs på grund av, då i N. och. excitation sker med hjälp av den sk. lokala strömmar (fig. 5).
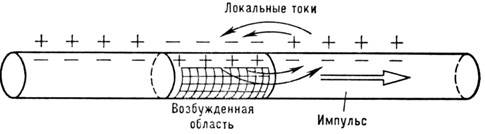
Ris. 5. Lokala strömmar ger distributionnervimpuls.
Ur-tion av Hodgkin - Huxley för N:s distribution och. löst numeriskt. De erhållna lösningarna tillsammans med de ackumulerade experimenten. uppgifter visade att N:s fördelning och. beror inte på detaljerna i excitationsprocessen. Kvaliteter. en bild av N:s utbredning och. kan erhållas med enkla modeller som endast reflekterar generella egenskaper upphetsning. Detta tillvägagångssätt gjorde det möjligt att beräkna hastigheten och formen på N. och. i en homogen fiber, deras förändring i närvaro av inhomogeniteter och till och med komplexa sätt för förökning av excitation i aktiva medier, till exempel. i hjärtmuskeln. Det finns flera matematik. modeller av detta slag. Den enklaste av dem är detta. Jonströmmen som flyter genom membranet under passagen av N. och. är teckenväxlande: först flyter den in i fibern och sedan ut. Därför kan den approximeras med en styckvis konstant funktion (fig. 2, G). Excitation uppstår när membranpotentialen förskjuts med tröskelvärdet j*. I detta ögonblick uppträder en ström, riktad inuti fibern och lika i absolut värde j". Efter en tid t "växlar strömmen till det motsatta, lika med j"". Denna fas fortsätter under en tid ~ t"". lösningen till ekvation (5) kan hittas som en funktion av variabeln t = x/ u, där u är fortplantningshastigheten för N. och. (Fig. 2, b).
I riktiga fibrer är tiden t" tillräckligt stor, så bara den bestämmer hastigheten u, för vilken f-la är giltig: ![]() . Givet att j" ~ ~d, R~d 2 och MED ~
d, Var d- fiberdiameter, finner vi i överensstämmelse med experimentet att u ~d 1/2. Med hjälp av en bitvis konstant approximation hittas formen på aktionspotentialen.
. Givet att j" ~ ~d, R~d 2 och MED ~
d, Var d- fiberdiameter, finner vi i överensstämmelse med experimentet att u ~d 1/2. Med hjälp av en bitvis konstant approximation hittas formen på aktionspotentialen.
Ur-tion (5) för spridning N. och. medger faktiskt två lösningar. Den andra lösningen visar sig vara instabil; det ger N. och. med mycket lägre hastighet och potentiell amplitud. Närvaron av den andra, instabila lösningen har en analogi i teorin om förbränning. När en låga fortplantar sig med en lateral kylfläns kan en instabil regim också uppstå. En enkel analys N:s modell och. kan förbättras med hänsyn till tilläggen. detaljer.
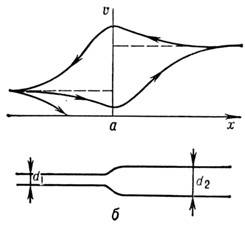
Vid snittbyte och vid förgrening av nervfibrer N:s passage och. kan vara svårt eller till och med helt blockerat. I en expanderande fiber (fig. 6) minskar pulshastigheten när den närmar sig expansion, och efter expansion börjar den öka tills den når ett nytt stationärt värde. N:s fördröjning och. desto starkare än mer skillnad i sektioner. När det räcker stor expansion N. och. stannar. Det finns en kritisk utvidgning av en fiber, ett snitt kvarhåller N. och.
Vid returrörelsen av N. och. (från bred fiber till smal) det finns ingen blockering, utan hastighetsförändringen är den motsatta. Vid ansatsen till att minska N:s hastighet och. ökar och börjar sedan falla till ett nytt stationärt värde. På hastighetsdiagrammet (fig. 6 A) visar sig vara en slags loop.
Rie. 6. Passage av nervimpulser genom att expanderalöpande fiber: A- förändring i hastigheten på impulsen in beroende på dess riktning; b- schematiskt expanderande fiberbild.
En annan typ av heterogenitet är fiberförgrening. I grennoden, olika alternativ för att skicka och blockera impulser. Vid den osynkrona N:s ansats och. blockeringsvillkoret beror på tidsförskjutningen. Om tidsförskjutningen mellan pulserna är liten hjälper de varandra att tränga in i den breda tredje fibern. Om skiftet är tillräckligt stort, så N. och. störa varandra. Detta beror på det faktum att N. och., som kom först, men misslyckades med att excitera den tredje fibern, delvis överför noden till ett eldfast tillstånd. Dessutom finns det en synkroniseringseffekt: i färd med N:s tillvägagångssätt och. till noden minskar deras fördröjning i förhållande till varandra.
N:s samspel och. Nervfibrer i kroppen kombineras till buntar eller nervstammar och bildar en sorts tvinnad kabel. Alla fibrer i ett knippe är oberoende. kommunikationslinjer, men har en gemensam "tråd" - den intercellulära vätskan. När N. och. löper längs någon av fibrerna, skapar det in interstitiell vätska elektrisk fältet, påverkar ett skär membranpotentialen hos angränsande fibrer. Vanligtvis är en sådan påverkan försumbar och kommunikationslinjerna fungerar utan ömsesidig störning, men det manifesterar sig i det patologiska. och konst. betingelser. Bearbetning nervstammar speciell. chem. ämnen, är det möjligt att observera inte bara ömsesidig interferens, utan också överföringen av excitation till närliggande fibrer.
Kända experiment på växelverkan mellan två nervfibrer placerade i en begränsad volym av yttre. lösning. Om N. löper längs med en av fibrerna och., så ändras excitabiliteten för den andra fibern samtidigt. Förändring går genom tre steg. Till en början sjunker excitabiliteten hos den andra fibern (excitationströskeln stiger). Denna minskning i excitabilitet föregår aktionspotentialen som rör sig längs den första fibern och varar ungefär tills potentialen i den första fibern når sitt maximum. Sedan växer excitabiliteten, detta stadium sammanfaller i tid med processen att minska potentialen i den första fibern. Excitabiliteten minskar igen när en lätt hyperpolarisering av membranet sker i den första fibern.
På samma gång N:s gång och. på två fibrer var det ibland möjligt att uppnå deras synkronisering. Trots att egen N:s hastigheter och. i olika fibrer är olika, samtidigt. excitation kunde uppstå kollektiva N. och. Om egen. hastigheterna var desamma, då hade den kollektiva impulsen lägre hastighet. Med en märkbar skillnad i egenskap. hastigheter hade den samlade hastigheten ett mellanvärde. Endast N. och. kunde synkronisera, vars hastigheter inte skilde sig alltför mycket.
Matta. beskrivningen av detta fenomen ges av ekvationssystemet för membranpotentialerna för två parallella fibrer j 1 och j 2:
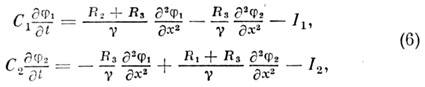
Var R 1 och R 2 - längsgående motstånd för de första och andra fibrerna, R 3 - längsgående motstånd yttre miljön, g = R 1 R 2 + R 1 R 3
+ R 2 R 3 . Jonströmmar jag 1 och jag 2 kan beskrivas med en eller annan modell av nervös excitation.
När du använder en enkel analys modelllösning leder till följande. bild. När en fiber exciteras, induceras en alternerande membranpotential i den intilliggande: först hyperpolariseras fibern, depolariseras sedan och till sist hyperpolariseras den igen. Dessa tre faser motsvarar en minskning, en ökning och en ny minskning av fiberns excitabilitet. Vid normala värden för parametrarna når skiftningen av membranpotentialen i den andra fasen mot depolarisering inte tröskeln, så det finns ingen överföring av excitation till den intilliggande fibern. På samma gång excitation av två fibrer, system (6) tillåter en självliknande gemensam lösning, vilket motsvarar att två N. och. rör sig med samma hastighet per stolpe. avstånd från varandra. Om det finns ett långsamt N. och. framåt, så saktar det ner den snabba impulsen, utan att släppa den framåt; båda rör sig i relativt låg hastighet. Om det finns en snabb II framåt. och., sedan drar den upp en långsam impuls. Den kollektiva hastigheten visar sig vara nära den inneboende hastigheten. snabb impulshastighet. I komplexa neurala strukturer, utseendet på auto kommer.
Spännande miljöer. Nervceller i kroppen kombineras till neurala nätverk, to-rye, beroende på frekvensen av förgrening av fibrerna, delas in i sällsynta och täta. I ett sällsynt nätverk element eldas oberoende av varandra och interagerar endast vid grennoder, som beskrivits ovan.
I ett tätt nätverk täcker excitationen många element samtidigt, så att deras detaljerade struktur och sättet de är sammankopplade visar sig vara obetydliga. Nätverket beter sig som ett kontinuerligt exciterbart medium, vars parametrar bestämmer förekomsten och fortplantningen av excitation.
Det exciterbara mediet kan vara tredimensionellt, även om det oftare betraktas som en tvådimensionell yta. Spänningen som uppstod i to-l. punkt på ytan, fortplantar sig i alla riktningar i form av en ringformad våg. Excitationsvågen kan gå runt hinder, men kan inte reflekteras från dem, inte heller reflekteras den från mediets gräns. När vågor kolliderar med varandra inträffar deras ömsesidiga förintelse; dessa vågor kan inte passera genom varandra på grund av närvaron av ett eldfast område bakom excitationsfronten.
Ett exempel på en excitabel miljö är hjärt neuromuskulärt syncytium - föreningen av nerv- och muskelfibrer till ett enda ledande system som kan överföra excitation i vilken riktning som helst. Neuromuskulär syncytia drar ihop sig synkront och lyder en våg av excitation, som skickas av ett enda kontrollcenter - pacemakern. En enda rytm störs ibland, arytmier uppstår. Ett av dessa lägen kallas förmaksfladder: dessa är autonoma sammandragningar som orsakas av cirkulationen av excitation runt ett hinder, till exempel. topp eller sämre ven. För förekomsten av en sådan regim måste omkretsen av hindret överstiga excitationsvåglängden, som är ~ 5 cm i det mänskliga förmaket. förmakskontraktion med en frekvens på 3-5 Hz. Ett mer komplext excitationsläge är flimmer. ventrikulärt hjärta, när odd. delar av hjärtmuskeln börjar dra ihop sig utan yttre. kommandon och utan kommunikation med angränsande element med en frekvens på ~ 10 Hz. Flimmer leder till att blodcirkulationen upphör.
Uppkomsten och upprätthållandet av spontan aktivitet hos ett exciterbart medium är oupplösligt kopplade till uppkomsten av vågkällor. Den enklaste källan till vågor (en grupp spontant exciterade celler) kan ge periodiska. pulsering av aktivitet, så fungerar hjärtats pacemaker.
Källor till excitation kan också uppstå på grund av komplexa utrymmen. organisering av excitationsläget, till exempel. efterklang av typen av en roterande spiralvåg, som uppträder i det enklaste exciterbara mediet. En annan typ av reverb förekommer i en miljö som består av två typer av element med olika excitationströsklar; reverbet exciterar periodiskt det ena eller andra elementet, samtidigt som det ändrar riktningen för dess rörelse och genererar.
Den tredje typen av källa är det ledande centret (ekokällan), som uppträder i en miljö som är inhomogen när det gäller eldfasthet eller excitationströskel. I detta fall uppträder en reflekterad våg (eko) på inhomogeniteten. Närvaron av sådana vågkällor leder till uppkomsten av komplexa excitationsregimer, som studeras i teorin om autovågor.
Belyst.: Hodgkin A., nervimpuls, per. från English, M., 1965; Katz B., Nerv, muskel och synaps, trans. från English, M., 1968; Khodorov B. I., The problem of excitability, L., 1969; Tasaki I., Nervös spänning, övers. från English, M., 1971; V. S. Markin, V. F. Pastushenko, Yu. A. Chizmadzhev, Theory of excitable media, Moskva, 1981. V. S. Markin.
Studiet av nervimpulsens natur var förknippat med särskilda svårigheter, eftersom det under impulsens passage längs nerven inte finns några synliga förändringar händer inte. Först nyligen, med utvecklingen av mikrokemiska metoder, var det möjligt att visa att under ledning av en impuls förbrukar nerven mer energi, förbrukar mer syre och frigör mer koldioxid än i vila. Detta indikerar att oxidativa reaktioner är involverade i ledningen av en impuls, i återställandet av det initiala tillståndet efter ledning, eller i båda dessa processer.
När man för ca 100 år sedan konstaterade att en nervimpuls åtföljs av vissa elektriska fenomen uppstod uppfattningen att själva impulsen är elektricitet. Vid den tiden var det känt att elektrisk ström färdades mycket snabbt, och därför uttrycktes åsikten att utbredningshastigheten för en nervimpuls var för stor för att kunna mätas. Tio år senare mätte Helmholtz hastigheten på impulsledning genom att stimulera en nerv som leder till en muskel för olika avstånd från muskeln och mäter tiden mellan irritation och sammandragning. På så sätt visade han att en nervimpuls fortplantar sig mycket långsammare än en elektrisk - i nerverna på en groda med en hastighet av cirka 30 m/s. Detta tydde naturligtvis på att nervimpulsen inte var en elektrisk ström, som strömmen i en koppartråd. Dessutom leder en död eller krossad nerv fortfarande ström men leder inte nervimpulser, och oavsett om vi irriterar nerven med ström, beröring, värme eller kemiska faktorer, impulsen som uppstår i detta fall fortplantar sig "I med en hastighet av samma ordning. Av detta drar vi slutsatsen att nervimpulsen inte är en elektrisk ström, utan en elektrokemisk störning i nervfibern. Störningen som orsakas av stimulansen i en sektion av nervfibern orsakar samma störning i nästa avsnitt, och så vidare tills impulsen når änden av fibern. Transmissionen av impulsen liknar således bränningen av en Fickford-sladd: från värmen som frigörs under förbränningen av en sektion av sladden, nästa sektion lyser etc. I nerven spelas värmens roll av elektriska fenomen som, efter att ha uppstått i ett område, stimulerar nästa.
Överföringen av en nervimpuls liknar bränningen av en säkring i vissa andra avseenden. En sladds brinnhastighet beror inte på mängden värme som går åt för att antända den, så länge den värmen är tillräcklig för att få sladden att fatta eld. Tändningsmetoden spelar ingen roll heller. Samma sak gäller med nerven. Nerven kommer inte att reagera förrän en stimulans av en viss minimistyrka appliceras på den, men ytterligare ökning av stimulans styrka kommer inte att orsaka att impulsen sprids snabbare. Detta beror på det faktum att energin för att leda impulsen tillförs av nerven själv och inte av stimulansen. Det beskrivna fenomenet återspeglas i allt-eller-inget-lagen: en nervimpuls är inte beroende av arten och styrkan hos stimulansen som orsakade den, om bara stimulansen är tillräckligt stark för att få impulsen att uppstå. Även om ledningshastigheten inte beror på stimulansens styrka, beror den på nervfiberns tillstånd och olika ämnen kan bromsa överföringen av en impuls eller göra det omöjligt.
En bränd sladd kan inte återanvändas, samtidigt som nervfibern kan återställa sitt ursprungliga tillstånd och överföra andra impulser. Den kan dock inte leda dem kontinuerligt: efter att ha genomfört en impuls går den över särskild tid innan fibern kan sända den andra pulsen. Denna tidsperiod, som kallas refraktärperioden, varar från 0,0005 till 0,002 sekunder. Under denna tid, kemiska och fysiska förändringar, som ett resultat av vilket fibern återgår till sitt ursprungliga tillstånd.
Så vitt vi vet är de impulser som överförs av neuroner av alla slag - motoriska, sensoriska eller interkalära - i grunden lika varandra. Den
en impuls orsakar en känsla av ljus, en annan en känsla av ljud, en tredje en muskelsammandragning och en fjärde stimulerar körtelns sekretoriska aktivitet, beror helt på naturen hos de strukturer till vilka impulserna kommer, och inte på några egenskaper av själva impulserna.
Även om en nervfiber kan stimuleras när som helst, under normala förhållanden, produceras stimulering endast i ena änden, från vilken impulsen färdas längs fibern till dess andra ände. Kopplingen mellan på varandra följande neuroner kallas synaps. En nervimpuls överförs från axonspetsen på en neuron till nästa dendrit genom en synaptisk korsning genom att en viss substans frigörs vid axonspetsen. Detta ämne orsakar uppkomsten av en nervimpuls i dendriten i nästa axon. Överföringen av excitation genom synapsen är mycket långsammare än dess överföring längs nerven. Under normala förhållanden färdas impulser endast i en riktning: i sensoriska neuroner går de från sinnesorganen till ryggmärgen och hjärnan, och i motorneuroner går de från hjärnan och ryggmärgen till musklerna och körtlarna. Riktningen bestäms av synapsen, eftersom endast spetsen av axonet kan frigöra ett ämne som stimulerar en annan neuron. Varje enskild nervfiber kan leda en impuls i båda riktningarna; när fibern stimuleras elektriskt någonstans i mitten uppstår två impulser, varav den ena går åt ena hållet och den andra åt den andra (dessa impulser kan detekteras av lämpliga elektriska anordningar). men bara den som går mot spetsen av axonet kan stimulera nästa neuron i kedjan. Impulsen som går till dendriten kommer att "stoppa" när den når slutet.
De kemiska och elektriska processer som är förknippade med överföringen av en nervimpuls liknar på många sätt de processer som sker under muskelkontraktion. Men den impulsledande nerven förbrukar väldigt lite energi jämfört med en sammandragande muskel; värmen som genereras under nervirritation under 1 minut, per 1 g vävnad, motsvarar den energi som frigörs under oxidationen av 0,000001 g glykogen. Detta "betyder att om nerven endast innehöll 1% glykogen som energikälla, kunde den stimuleras kontinuerligt i en vecka och glykogentillförseln skulle inte vara slut. Med tillräcklig tillförsel av syre är nervfibrerna praktiskt taget outtröttliga. naturen" mental trötthet", detta kan inte vara en verklig trötthet av nervtrådarna.
Acetylkolins roll
Den neuromuskulära korsningen har studerats i detalj, och detta har gett mycket information användbar för att förstå processen för kemisk överföring i neuronala synapser i allmänhet. Dess funktion är att överföra impulser från relativt mycket små motoriska nervändar till en stor muskelfiber och få den att dra ihop sig. I de flesta myoneurala knutpunkter i ryggradsdjursmuskler följs varje nervimpuls av samma impuls i muskelfibrerna, som snabbt fortplantar sig i båda riktningarna mot senorna, vilket ger tillräckligt synkron aktivering av kontraktila proteiner från ena änden av fibern till den andra. Den myoneurala korsningen hos ryggradsdjur tjänar alltså ett mycket enklare syfte än neuronernas centrala synapser eller de perifera excitatoriska och hämmande neuromuskulära korsningarna hos kräftdjur, där integrationen av konvergenta signaler sker och där effekten av en enskild nervimpuls vanligtvis ligger långt under excitationen. effektorcellens tröskel. Grovt sett spelar ryggradsdjurets neuromuskulära korsning rollen som ett enkelt relä. Den kan också ses som en impedansmatchningsanordning, en sorts pulsomvandlare som matar ut en elektrisk ström som är tillräcklig för att höja potentialen hos ett muskelmembran med låg impedans över en tröskel.
Som redan noterats, på basis av enbart strukturella data, kan det sägas att förekomsten av en effektiv elektrisk kabelförbindelse mellan nerven och muskeln är extremt osannolik. I grodans skelettmuskel bryts den myeliniserade nervfibern upp i en hel "buske" av terminala icke-köttiga grenar ca 1,5 mikron tjocka, som passerar i små skåror på muskelfiberns yta i ca 100 mikron. Genom hela denna terminala sektion bildar nerven en synaptisk förbindelse med muskelfibern. På många ställen inne i nervändarna låter elektronmikroskopet oss se de vanliga ansamlingarna av bubblor med en diameter på 500 A (fig. 27). De presynaptiska och postsynaptiska membranen är åtskilda av ett intercellulärt utrymme som innehåller ett basalmembran. Ytan av muskelfibern bildar en regelbunden rad av veck som ligger i rät vinkel mot nervens terminala gren.
Låt oss se vilka möjligheter som finns för elektrisk impulsöverföring genom en typisk myoneural junction i en av de tjockaste grodmuskelfibrerna (150 mikrometer i diameter). För en kort strömpuls, såsom den som produceras av en nervaktionspotential, är motståndet (ingångsimpedansen) hos muskelfibern mindre än 50 000 ohm. För att excitera en fiber måste dess vilande membranpotential sänkas från 90 mV till cirka 50 mV, och detta kräver en strömpuls på något mindre än 10-6 A. (Senare kommer vi att se att kemisk överföring genererar en ström på 2-3-10"6 ampere i många ändplattor, och därför har processen gott om "reservkraft".)
Vilken strömstyrka kan nervändarna ge under elektrisk överföring? Vi kommer att anta det total längd/ synaptiska förgreningar av nerven vid ena ändplattan är cirka 1 mm, och den "synaptiska ytan" (mot muskelfibern) beräknas med formeln ir / w 2,3-10 "5 cm2. Om nervmembranet skapar en utåtgående ström puls med en densitet i storleksordningen 1 A" cm2 (det vanliga värdet för en icke-myelinös nerv och för en muskel), kommer detta att ge en total ström på 2,3 x 10"8 A. Även om all denna ström gick in i muskelfibern i det synaptiska området (vilket kan hända i fallet med kontinuitet av protoplasma mellan nerv och muskel) och därmed kunde depolarisera muskelmembranet, så kan det även då bara förändra potentialen: "
"¦Nervös ¦Shnchaiye
)~~Tmk" "^Muskulär i / yukno";
muskelfiber
Schwann bur
nervändan
Fikon. 27. Schema av synaptiska strukturer (visad skala är endast ungefärlig),
I. Interneuronal synaps: A - struktur av den presynaptiska ändelsen; B - synapsens placering på neuronens kropp. II. Neuromuskulär grodanslutning: A - separat del av anslutningen; B - den allmänna platsen för ändarna av motoraxonen på muskelfiber(ytan som visas i diagram A indikeras av en liten rektangel); B - en schematisk ritning gjord från ett elektronmikrofoto av ett längsgående snitt av en muskelfiber: I - axonterminalmembran; 2 - "basalmembran", som ligger i gapet mellan nerv- och muskelfibrerna; 3-vikt postsynaptiskt membran av muskelfibern.
för 1-2 mv. Men i verkligheten, på grund av närvaron av ett intercellulärt gap som är mer än 500 A brett och impedansen hos det postsynaptiska membranet (fig. 26 och 27), är förutsättningarna för strömutbredning mycket sämre. Om spaltens resistivitet har ett värde i storleksordningen 100 ohm-cm, blir den potentiella förändringen i den endast 40 mikrovolt. Då kommer strömmen som kommer in i muskelfibern att försvagas till ett värde i storleksordningen 10-11 A och kommer att kunna ändra sin membranpotential endast med en obetydlig mängd (mindre än 1 mikrovolt).
Trots den mycket trevande karaktären hos ovanstående beräkningar är det tydligt att bristen på strukturell kontinuitet i den synaps vi har ansett gör elkabelöverföring extremt osannolik; impedanserna i nervändarna och muskelfibern är så "felmatchade" att även om cytoplasman var kontinuerlig, skulle det vara svårt att föreställa sig ett system som är mindre lämpat för elektrisk signalering!
Flera försök har gjorts att testa detta i ett ex-experiment. Om en undertröskelström appliceras på en nerv nära dess synaps, orsakar den ingen märkbar förändring av den lokala potentialen i muskelfibern (se även s. 169). När en impuls uppstår fortplantar den sig till ändarna av omyeliniserade grenar och exciterar en impuls i muskelfibern. Om mikroelektroden placeras exakt mot den synaptiska kontakten mellan nerv och muskel, separeras två elektriska förändringar(Fig. 28): först en elektrisk ström genererad av en impuls i axonets terminala förgrening, och sedan, efter 0,5-0,8 meeks, en annan liknande ström som uppstår i det postsynaptiska membranet. Dessa två händelser är inte bara åtskilda i tid; den postsynaptiska förändringen kan också tas bort selektivt, antingen med hjälp av vissa ämnen, såsom curare, eller genom att sänka koncentrationen av kalcium eller öka koncentrationen av magnesium. Den observerade fördröjningen på platserna för myoneural kontakt är ett konstant och mycket viktigt fenomen: det indikerar tydligt frånvaron av elektrisk kontinuitet i synapsen. Uppenbarligen kabelprocessen i presynaptiska
muskelfiber
r-0-
Artefaktimpuls,
irritation kommer
Jag är ungefär nervös
jag slutar
3
І -
11111111
Tid, ms »¦
Fikon. 28. Punktabduktion från ytan av den neuromuskulära korsningen.
Med brist på kalcium och ökad koncentration magnesium postsynaptisk avvikelse kan saknas ("noll" reaktion 1) eller uppträda som en "kvant" reaktion 2. Under normala förhållanden finns det en mycket större avvikelse 3, vilket leder till muskelspikar.
slutet stannar, och någon annan, icke-elektrisk process fungerar som en mellanlänk mellan ankomsten av en nervimpuls och uppkomsten av en elektrisk signal i muskelfibern.
Den kemiska naturen hos denna mellanprocess avslöjades i Sir Henry Dales och hans medarbetares arbete. , som visade det som ett resultat av irritation motorisk nerv acetylkolin frisätts och att detta ämne har en mycket stark stimulerande effekt på skelettmuskulaturen. För att fastställa acetylkolins roll som en kemisk transmitter, eller mediator, var det nödvändigt att visa: 1) att det frisätts från något presynaptiskt ställe, 2) att det verkar på något postsynaptiskt ställe, och 3) att mängden av det som utsöndras av nerven är tillräckligt för att initiera en impuls i muskeln.
mänskligt nervsystem
Frågor med val av ett rätt svar.
A1. Nervös reglering av funktioner i människokroppen utförs med hjälp av:
1) elektriska impulser,
2) mekanisk irritation,
3) hormoner,
4) enzymer.
A2. Strukturella och funktionell enhet nervsystemet överväga:
1) neuron,
2) nervvävnad,
3) ganglioner,
4) nerver.
A3. grund nervös aktivitet människa och djur är:
1) tänka,
2) rationell aktivitet,
3) spänning,
4) reflex.
A4. Receptorer är känsliga strukturer som:
1) överföra impulser till centrala nervsystemet,
2) överför nervimpulser från interkalära neuroner till verkställande,
3) uppfatta stimuli och omvandla stimulans energi till en process av nervös excitation,
4) uppfatta nervimpulser från känsliga neuroner.
A5. Celler som är mest känsliga för brist på syre:
1) ryggmärgen,
2) hjärnan,
3) lever och njurar,
4) mage och tarmar.
A6. Buntar av långa processer av neuroner, täckta med en bindvävsskida och placerade utanför det centrala nervsystemet, bildar:
1) nerver,
2) lillhjärnan,
3) ryggrad,
4) skällande halvklot.
A7. Frivilliga mänskliga rörelser ger:
1) lillhjärnan och diencephalon,
2) mitten och ryggmärgen,
3) märg och bro,
4) stora hemisfärer i framhjärnan.
A8. Reglering och harmonisering fysiologiska processer flödar i de inre organen, ger:
1) diencephalon,
2) mellanhjärnan,
3) ryggmärgen,
4) lillhjärnan.
A9. Det somatiska nervsystemet, till skillnad från det autonoma nervsystemet, styr arbetet med:
1) skelettmuskel,
2) hjärta och blodkärl,
3) tarmar,
4) njurar.
A10. Nervimpulser överförs till hjärnan genom neuroner:
1) motor,
2) infoga,
3) känslig,
4) verkställande.
A11. Centren för sväljnings-, andnings-, kardiovaskulära och andra vitala reflexer är belägna i:
1) lillhjärnan,
2) mellanhjärnan,
3) medulla oblongata,
4) diencephalon.
A12. Det autonoma nervsystemet är involverat i:
1) genomförandet av godtyckliga rörelser,
2) uppfattning om visuella, auditiva och smakstimuli,
3) reglering av ämnesomsättning och arbete inre organ,
4) bildandet av talljud.
A13. En nervimpuls kallas:
1) en elektrisk våg som färdas längs en nervfiber,
2) överföring av information från en neuron till nästa,
3) överföring av information från cell till cell,
4) en process som ger hämning av målcellen.
A14. Excitation riktas längs den känsliga neuronen:
1) till det centrala nervsystemet,
2) till verkställande organ,
3) till receptorer,
4) till musklerna.
A15. Nervimpulser överförs från sinnesorganen till hjärnan via:
1) motoriska neuroner,
2) interkalära neuroner,
3) känsliga neuroner,
4) korta processer av motorneuroner.
A16. Externa stimuli omvandlas till nervimpulser i:
1) nervfibrer,
2) kroppar av nervceller i det centrala nervsystemet,
3) receptorer,
4) kroppar av interkalära neuroner.
A17. Hos människor är pupillvidgning ansvarig för:
1) sympatisk avdelning nervsystem,
2) parasympatisk uppdelning nervsystem,
4) centrala nervsystemet.
A18. Den korta förlängningen av en nervcell kallas:
1) axon,
2) neuron,
3) dendrit,
4) synaps.
A19. Den långa förlängningen av en nervcell kallas:
1) axon,
2) neuron,
3) dendrit,
4) synaps.
A20. Punkten där två nervceller möter varandra kallas:
1) axon,
2) neuron,
3) dendrit,
4) synaps.
A21. Nerver är:
1) neural krets,
2) ansamling av nervceller,
3) knippen av axoner som sträcker sig utanför hjärnan,
4) receptorer.
A22. Hjärnhalvorna är anslutna till varandra:
1) bro,
2) corpus callosum,
3) mellanhjärnan,
4) diencephalon.
A23. Påverkan av det parasympatiska nervsystemet på hjärtaktiviteten uttrycks i:
1) långsam hjärtslag,
2) ökad hjärtfrekvens,
3) hjärtstopp,
4) arytmier.
A24. Nervsystemet är:
1) kropp,
2) tyg,
3) organsystem,
4) organoid.
A25. Det mänskliga nervsystemet, i motsats till det endokrina:
1) svarar på yttre, inte på inre påverkan,
2) helt underställd medvetande,
3) agerar snabbare
4) fungerar inte under sömnen.
A26. Reflexer som inte kan förstärkas eller hämmas av en persons vilja utförs genom nervsystemet:
1) central,
2) vegetativ,
3) somatisk,
4) perifer.
A27. Axoner - processer av nervceller som sträcker sig utanför det centrala nervsystemet, samlas i buntar och bildar:
1) subkortikala kärnor,
2) nervknutor,
3) cerebellar cortex,
4) nerver.
A28. Neuron är:
1) en multinukleär cell med processer,
2) en enkärnig cell med processer,
3) en kärnkraftsfri cell med processer,
4) en multinukleär cell med cilia.
A29. I kroppens adaptiva reaktioner på förändringar i miljöförhållanden spelas den ledande rollen av:
1) hjärna,
2) autonoma nervsystemet,
3) somatiska nervsystemet,
4) sinnesorgan.
A30. Nervceller skiljer sig från andra genom att ha:
1) kärnor med kromosomer,
2) processer olika längder,
3) flerkärniga,
4) kontraktilitet.
A31. Överföringen av excitation längs en nerv eller muskel förklaras av:
1) skillnaden i koncentrationerna av natrium- och kaliumjoner inuti och utanför cellen,
2) bryta vätebindningar mellan vattenmolekyler,
3) en förändring i koncentrationen av vätejoner,
4) vattens värmeledningsförmåga.
A32. Reflex, nervcentrum som ligger utanför medulla oblongata:
1) hosta,
2) svälja,
3) salivutsöndring,
4) knä.
A33. Diencephalon reglerar:
1) ämnesomsättning,
2) mat och vattenintag,
3) underhåll konstant temperatur kropp,
4) alla svar är korrekta.
A34. Reflexens centrum är beläget i medulla oblongata:
1) nysningar,
2) urinering,
3) avföring,
4) knä.
A35. Host- och nysningscenter finns i:
1) ryggmärgen,
2) medulla oblongata,
3) mellanhjärnan,
4) framhjärna.
A36. Det parasympatiska nervsystemet minskar:
1) frekvens hjärtfrekvens,
2) kraften av hjärtsammandragningar,
3) plasmaglukosnivå,
4) alla listade parametrar.
Frågor med val av flera rätta svar.
I 1. vit substans främre sektionen hjärna:
A) bildar dess bark,
B) ligger under barken,
B) består av nervfibrer
D) bildar subkortikala kärnor,
D) förbinder hjärnbarken med andra delar av hjärnan och med ryggmärgen,
E) utför funktionen av en högre analysator av signaler från alla receptorer i kroppen.
AT 2. Vilka organ regleras av människans autonoma nervsystem?
A) muskler i övre och nedre extremiteterna,
B) hjärta och blodkärl
I) matsmältningsorgan,
G) ansiktsmuskler,
D) njurar och Blåsa,
E) diafragma och interkostala muskler.
VID 3. Det perifera nervsystemet inkluderar:
A) en bro
B) lillhjärnan
B) nervknutor
D) ryggmärg
D) känselnerver,
E) motoriska nerver.
AT 4. I cerebellum finns regleringscentra:
A) muskeltonus
B) vaskulär tonus,
B) poser och kroppsbalans,
G) rörelsekoordination,
D) känslor
E) andas in och andas ut.
Efterlevnadsuppgifter.
VID 5. Upprätta en överensstämmelse mellan en viss funktion hos en neuron och den typ av neuron som utför denna funktion.
VID 6. Upprätta en överensstämmelse mellan nervsystemets delar och deras funktioner.
| UTFÖRDA FUNKTIONER | AVDELNING FÖR NERVSYSTEMET |
| 1) drar ihop blodkärlen, | A) söt |
| 2) saktar ner rytmen hjärtats verk, | B) parasympatisk. |
| 3) smalnar av bronkerna, | |
| 4) vidgar pupillen. |
KLOCKAN 7. Upprätta en överensstämmelse mellan strukturen och funktionerna hos en neuron och dess processer.
VID 8. Upprätta en överensstämmelse mellan nervsystemets egenskaper och dess typer som har dessa egenskaper.
VID 9. Upprätta en överensstämmelse mellan exempel på mänsklig nervös aktivitet och ryggmärgens funktioner.
KLOCKAN 10. Upprätta en överensstämmelse mellan de strukturella egenskaperna och funktionerna i hjärnan och dess avdelning.
Uppgiften att fastställa rätt sekvens.
VID 11. Installera rätt sekvens placeringen av delarna av hjärnstammen, i riktning från ryggmärgen.
A) diencephalon
B) medulla oblongata
B) mellanhjärnan
D) bro.
Gratis svarsfrågor.
C1. Hitta fel i den givna texten. Ange antalet meningar som gjorde fel, förklara dem.
1. Hjärnbarken bildas av grå substans.
2. grå materia består av processer av neuroner.
3. Varje hemisfär är uppdelad i frontal-, parietal-, temporal- och occipitallober.
4. Synzonen är belägen i frontalloben.
5. Den auditiva zonen är belägen i parietalloben.
C2. Hitta fel i den givna texten. Ange antalet meningar där fel gjordes, förklara dem.
1. Nervsystemet är uppdelat i centrala och somatiska.
2. Det somatiska nervsystemet är uppdelat i perifert och autonomt.
3. Centralavdelning Det somatiska nervsystemet består av ryggmärgen och hjärnan.
4. Det autonoma nervsystemet koordinerar skelettmusklernas aktivitet och ger känslighet.
Svar på uppgifterna i del A.
| № | ||||||||||||
| svar | ||||||||||||
| № | ||||||||||||
| svar | ||||||||||||
| № | ||||||||||||
| svar |
Svar på uppgifterna i del B med val av flera rätta svar.
| Jobb nummer | ||||||
| svar | ||||||
| Jobb nummer | ||||||
| svar |
Liknande artiklar
-
När en man är emot ett barn, hur blir man gravid utan hans vetskap?
Ibland kan man bli gravid av oaktsamhet. För att förhindra att detta inträffar är det viktigt att veta hur du kan bli gravid med ett barn av misstag och vilka medel du kan använda för att undvika oönskad graviditet. Även i den här artikeln kan du hitta information om ...
-
Vilka stenar och amuletter är lämpliga för Oxen enligt horoskopet och födelsedatumet Elefanttalisman för Oxen
April-maj Oxen (21 april - 20 maj) är mätt, inte kinkig och kolossalt produktiv! Deras avundsvärda envishet kan få andra att ta tag i, men de vet exakt vad de gör och varför de behöver det. Bland de positiva...
-
Restriktioner för åtkomst till data i roller 1c
Alla användarrättighetsinställningar som vi kommer att göra inom ramen för denna artikel finns i avsnitt 1C 8.3 "Administration" - "Användar- och rättighetersinställningar". Denna algoritm är liknande i de flesta konfigurationer på ...
-
1c startar en tunn klient istället för en tjock
Plattformar: 1C: Enterprise 8.3, 1C: Enterprise 8.2, 1C: Enterprise 8.1 Konfigurationer: Alla konfigurationer2012-11-16 21362 De lanseras genom att specificera speciella ...
-
Bevis på kända sätt att stjäla el Hur man tar reda på vem som stjäl el
Höjningen av energitaxor är ett av de slående dragen i den fördjupade ekonomiska krisen. I detta sammanhang är stöld av el och de problem som är förknippade med dess upptäckt av största vikt. Sätt att upptäcka stöld ...
-
Funktioner för montering av uttag och strömbrytare på olika ytor
Hälsningar till alla läsare av vår blogg Idag, kära läsare, vill jag lyfta fram ämnet om hur man installerar uttag. Denna procedur efterfrågas mycket ofta när man byter ut ett gammalt uttag med ett nytt i händelse av ett haveri, när ...