Naučnici su izbrojali broj neurona u ljudskom mozgu. Električne manifestacije kortikalne aktivnosti
Kora hemisfera veliki mozak predstavlja najviše i najsloženije organizovano nervnog centra tipa ekrana, čija aktivnost osigurava regulaciju različitih tjelesnih funkcija i složenih oblika ponašanja.
Korteks je formiran slojem sive tvari debljine 3-5 mm. Siva tvar sadrži nervne ćelije (više od 10 milijardi neurona), nervna vlakna i neuroglijalne ćelije. Njegovi različiti dijelovi, koji se međusobno razlikuju po određenim karakteristikama položaja i strukture ćelija, rasporedu vlakana i funkcionalnom značaju, nazivaju se poljima, koje je prvi opisao njemački liječnik i znanstvenik K. Brodmann.
Citoarchitecture
Među multipolarnim neuronima korteksa razlikuju se piramidalne, zvjezdaste, vretenaste, arahnidne, horizontalne, "kandelabraste" stanice, stanice s dvostrukim buketom dendrita i neke druge vrste neurona.
Piramidalni neuroničine osnovni i najspecifičniji oblik za moždanu koru. Imaju izduženo tijelo u obliku konusa, čiji je vrh okrenut prema površini korteksa. Dendriti se protežu od vrha i bočnih površina tijela. Aksoni potiču iz baze piramidalnih ćelija.
Piramidalne ćelije različitih slojeva korteksa razlikuju se po veličini i razlikuju se funkcionalna vrijednost. Male ćelije su interneuroni. Aksoni velikih piramida učestvuju u formiranju motor piramidalne staze.
Neuroni korteksa nalaze se u nejasno omeđenim slojevima, koji su označeni rimskim brojevima i numerisani izvana prema unutra. Svaki sloj karakterizira prevlast jedne vrste ćelija. Postoji šest glavnih slojeva u moždanoj kori:
- I - molekularni;
- II - eksterno zrnasto;
- III - piramidalni;
- IV - unutrašnje zrnasto;
- V - ganglijski;
- VI - sloj polimorfnih ćelija.
ja - Molekularno sloj kore sadrži br veliki broj male asocijativne horizontalne ćelije Cajal. Njihovi aksoni idu paralelno s površinom mozga kao dio tangencijalnog pleksusa nervnih vlakana molekularnog sloja. Međutim, najveći dio vlakana ovog pleksusa predstavljen je grananjem dendrita donjih slojeva.
II - Eksterno granulirano sloj formiraju brojni mali piramidalni i zvjezdasti neuroni. Dendriti ovih ćelija uzdižu se u molekularni sloj, a aksoni ili ulaze u njih bijele tvari, ili, formirajući lukove, također ulaze u tangencijalni pleksus vlakana molekularnog sloja.
III - Najširi sloj kore velikog mozga - piramidalni. Sadrži piramidalne neurone, Martinottijeve ćelije i vretenaste ćelije sa dvostrukim buketom dendrita. Apikalni dendriti piramida protežu se u molekularni sloj, a lateralni dendriti formiraju sinapse sa susjednim ćelijama ovog sloja. Akson piramidalne ćelije uvijek se proteže od njene baze. U malim ćelijama ostaje unutar korteksa, u velikim ćelijama formira mijelinsko vlakno koje ide u bijelu tvar mozga. Aksoni malih poligonalnih Martinotti ćelija usmjereni su u molekularni sloj. Piramidalni sloj obavlja prvenstveno asocijativnu funkciju.
IV - Unutrašnja zrnasta sloj u nekim poljima korteksa je vrlo razvijen (na primjer, u vidnim i slušnim područjima korteksa), dok u drugim može biti gotovo odsutan (na primjer, u precentralnom girusu). Ovaj sloj formiraju mali zvjezdasti neuroni. Sadrži veliki broj horizontalnih vlakana.
V- Ganglijski sloj korteksa formiran je od velikih piramida, a područje motornog korteksa (precentralni girus) sadrži gigantske piramide , koje je prvi opisao kijevski anatom V. A. Bets. Apikalni dendriti piramida dosežu prvi sloj. Aksoni piramida projektuju se na motorna jezgra mozga i kičmena moždina. Najduži aksoni Betzovih ćelija u piramidalnim traktovima dosežu kaudalne segmente kičmene moždine.
Pored piramidalnih neurona, ganglijski sloj korteksa sadrži vertikalne ćelije u obliku vretena, čiji se aksoni uzdižu u 1. sloj korteksa, kao i ćelije košare.
VI - Sloj polimorfne ćelije formirani od neurona različitih oblika (fusiformne, zvjezdaste, Martinottijeve stanice). Aksoni ovih stanica protežu se u bijelu tvar kao dio eferentnih puteva, a dendriti dopiru do molekularnog sloja.
Myeloarchitecture
Među nervnim vlaknima moždane kore možemo razlikovati asocijativni vlakna koja povezuju pojedina područja korteksa jedne hemisfere, commissural, povezujući korteks različitih hemisfera, i projekcija vlakna, i aferentna i eferentna, koja povezuju korteks sa jezgrima nižih delova centralnog nervnog sistema. Projekciona vlakna u korteksu velikog mozga formiraju radijalne zrake koje završavaju u trećem piramidalnom sloju. Pored već opisanog tangencijalnog pleksusa I - molekularnog sloja, na nivou IV - unutrašnjeg granularnog i V - ganglijskog sloja nalaze se dva tangencijalna sloja mijelinskih nervnih vlakana - respektivno, spoljna traka Baillargera i unutrašnja traka Baillarger. Posljednja dva sistema su pleksusi formirani od završnih dijelova aferentnih vlakana.
Modularni princip organizacije korteksa
U moždanoj kori opisani su ponavljajući blokovi, odnosno moduli neurona, koji se smatraju njegovim morfofunkcionalnim jedinicama sposobnim za relativno autonomnu aktivnost. Imaju oblik cilindara, odnosno stupova, koji prolaze okomito kroz cijelu debljinu kore.
Svaki modul uključuje aferentne puteve, sistem lokalnih veza i eferentne puteve.
TO aferentnih puteva uključuju kortiko-kortikalna i talamo-kortikalna vlakna.
Modul je organiziran oko kortiko-kortikalnih vlakana, koji su aksoni piramidalnih stanica ili iste ili suprotne hemisfere. Kortiko-kortikalna vlakna formiraju završetke u svim slojevima korteksa datog modula.
Modul takođe uključuje talamo-kortikalna vlakna koja se završavaju u sloju IV korteksa na bodljastim zvezdastim neuronima i bazalnim dendritima piramidalnih neurona.
Eferentni putevi formiraju aksoni velikih i džinovskih piramidalnih neurona, kao i aksoni fusiformnih i nekih drugih ćelija sloja VI korteksa.
Sistem lokalnih veza formiraju interneuroni modula, koji uključuju više od desetak tipova ćelija. Većina njih je inhibitorna i reguliše aktivnost pretežno piramidalnih neurona.
Od inhibitornih neurona modula najveća vrijednost imati:
- akso-aksonske ćelije;
- ćelije "kandelabra";
- košaraste ćelije;
- ćelije sa dvostrukim buketom dendrita;
- ćelije aksonalnom četkicom.
Sistem inhibitornih neurona igra ulogu filtera, inhibirajući dio piramidalnih neurona korteksa.
Moždane opne mozga i kičmene moždine
Mozak i kičmena moždina prekriveni su sa tri vezivnotkivne membrane: mekom, neposredno uz moždano tkivo, arahnoidnom i tvrdom, koja se graniči sa koštanog tkiva lobanje i kičme. Moždane ovojnice pružaju zaštitu, uklj. funkcija apsorpcije udara, osigurava proizvodnju i apsorpciju likvora.
Pia mater neposredno uz moždano tkivo i ograničeno od njega marginalnom glijalnom membranom. U rastresitim vlaknima vezivno tkivo Ljuska sadrži veliki broj krvnih sudova koji opskrbljuju mozak, brojna nervna vlakna, terminalni aparat i pojedinačne nervne ćelije. Pia mater okružuje sudove koji prodiru u mozak, formirajući oko njih perivaskularnu pijalnu membranu. U komorama mozga, pia mater, zajedno s ependimom, sudjeluje u formiranju horoidnih pleksusa koji proizvode cerebrospinalnu tekućinu.
Arahnoidna predstavljen tankim slojem labavog vlaknastog vezivnog tkiva. Između nje i pia mater nalazi se mreža poprečnih šipki koja se sastoji od tankih snopova kolagenih i tankih elastičnih vlakana. Ova mreža povezuje školjke jedna s drugom. Između pia mater, koja prati reljef moždanog tkiva, i arahnoida, koji se proteže uz povišena područja ne ulazeći u udubljenja, nalazi se subarahnoidalni (subarahnoidalni) prostor, prožet tankim kolagenim i elastičnim vlaknima koja povezuju membrane sa svakim. ostalo. Subarahnoidalni prostor komunicira s komorama mozga i sadrži cerebrospinalnu tekućinu. U ovom prostoru postoje velike krvni sudovi, čije grane hrane mozak.
Resice arahnoidne membrane (najveće se nazivaju Pachyon granulacije) služe kao mjesta kroz koja se tvari iz cerebrospinalne tekućine vraćaju u krv. Oni su avaskularne izrasline arahnoidne membrane, koje sadrže mrežu proreznih prostora i strše u lumen sinusa dura mater.
Dura mater formirano od gustog vlaknastog vezivnog tkiva koje sadrži mnoga elastična vlakna. U kranijalnoj šupljini je čvrsto spojen sa periostom. IN kičmeni kanal dura mater je od periosteuma kralješka ograničena epiduralnim prostorom, ispunjenim slojem labavog vlaknastog vezivnog tkiva, što joj daje određenu pokretljivost. Između dura mater i arahnoidne membrane nalazi se subduralni prostor. Subduralni prostor sadrži malu količinu tečnosti.
Membrane sa strane subduralnog i subarahnoidalnog prostora prekrivene su slojem ravne ćelije glijalna priroda.
Promjene vezane za dob
Promjene u centralnom nervnom sistemu u starost prvenstveno povezan sa sklerotskim promjenama u krvnim žilama mozga. U starijoj dobi dolazi do zadebljanja pia mater i arahnoidalne membrane mozga. U njima se mogu pojaviti naslage kamenca. Uočava se kortikalna atrofija moždane hemisfere, prvenstveno frontalni i parijetalni režnjevi. Broj neurona po jedinici volumena korteksa se smanjuje, što uglavnom ovisi o smrti stanice. Neuroni se smanjuju u veličini, djelomično gube svoju bazofilnu tvar, jezgre postaju gušće, a njihov obris postaje neujednačen. Piramide sloja V motornog korteksa i piriformne ćelije kore malog mozga mijenjaju se brže od ostalih. Lipofuscin granule se akumuliraju u neuronima različitih dijelova nervnog sistema.
Sve najvažnije reakcije ponašanja ljudi odvijaju se uz pomoć centralnog nervnog sistema. Glavne funkcije centralnog nervnog sistema su:
- ujedinjenje svih delova tela u jedinstvenu celinu i njihovo regulisanje;
- upravljanje stanjem i ponašanjem tela u skladu sa uslovima spoljašnje okruženje i njegove potrebe.
Kod viših životinja i ljudi Vodeći dio centralnog nervnog sistema je moždana kora . On kontrolira najsloženije funkcije u ljudskom životu mentalnih procesa(svijest, mišljenje, govor, pamćenje, itd.).
Glavne metode za proučavanje funkcija centralnog nervnog sistema su metode uklanjanja i iritacije (u klinici i kod životinja), snimanje električnih pojava, metoda uslovljeni refleksi.
I dalje se razvijaju nove metode za proučavanje centralnog nervnog sistema: korišćenjem tzv kompjuterizovana tomografija možete vidjeti morfofunkcionalne promjene u mozgu na različitim dubinama; fotografija u infracrvenim zracima (termalna slika) omogućava vam da otkrijete „najtoplije“ tačke u mozgu; Nove podatke o funkcionisanju mozga daje proučavanje njegovih magnetnih oscilacija.
Osnovne funkcije i interakcije neurona
Glavni strukturni elementi nervnog sistema su nervne ćelije ili neuroni.
Osnovne funkcije neurona
Putem neurona, informacije se prenose s jednog dijela nervnog sistema na drugi, informacije se razmjenjuju između nervnog sistema i različitih dijelova tijela. Najsloženiji procesi obrade informacija odvijaju se u neuronima. Uz njihovu pomoć se formiraju odgovore tijela (refleksi) na vanjske i unutrašnje podražaje.
dakle, glavne funkcije neurona su: percepcija vanjskih podražaja funkciju receptora, njihovu obradu integrativna funkcija i prenošenje nervnih uticaja na druge neurone ili različite radne organe efektorska funkcija. Glavni procesi obrade informacija odvijaju se u tijelu nervne ćelije ili some. Brojni razgranati izdanci u obliku drveća dendriti(grčko dendron drvo) služe kao neuronski ulazi preko kojih signali ulaze u nervnu ćeliju. Izlaz neurona je proces koji se proteže od tijela ćelije akson(grč. axis axis), koja prenosi nervne impulse dalje do druge nervne ćelije ili radnog organa (mišića, žlezde). Početni dio aksona i njegov produžetak na mjestu njegovog izlaska iz tijela ćelije posebno su jako ekscitabilni. axon hilllock neuron. U ovom segmentu ćelije nastaje nervni impuls.
Vrste neurona
Neuroni se dijele na tri glavna tipa: aferentni, eferentni i intermedijarni. Aferentni neuroni(osjetljivi ili centripetalni) prenose informacije od receptora do 1CNS-a. Tijela ovih neurona nalaze se izvan CNS-a kičmeni čvorovi i na čvorovima kranijalni nervi. Aferentni neuroni imaju dug proces - dendrit, koji je na periferiji u kontaktu sa perceptivnom formacijom - receptor ili sam formira receptor, kao i drugi proces - akson, koji ulazi kroz zadnje rogove u kičmenu moždinu.
Eferentni neuroni(centrifugalni) su povezani sa prenošenjem silaznih uticaja sa gornjih podova nervnog sistema na one ispod ili sa centralnog nervnog sistema na radne organe. Eferentne neurone karakterizira razgranata mreža kratkih procesa - dendrita i jednog dugog procesa - aksona.
Srednji neuroni(interneuroni ili interneuroni) to su, po pravilu, manje ćelije koje komuniciraju između različitih (posebno aferentnih i eferentnih) neurona. Oni prenose nervne utjecaje u horizontalnom smjeru (na primjer, unutar jednog segmenta kičmene moždine) i u vertikalnom smjeru (na primjer, iz jednog segmenta kičmene moždine u drugi gornji ili donji segment). Zbog brojnih grana aksona, interneuroni mogu istovremeno pobuđivati veliki broj drugih neurona.
Ekscitatorne i inhibitorne sinapse
Interakcija neurona jedni s drugima (i sa efektornim organima) se odvija putem specijalno obrazovanje sinapse (grčki: kontakt). Nastaju od završnih grana neurona na tijelu ili od procesa drugog neurona. Što je više sinapsi na nervnoj ćeliji, ona više percipira razne iritacije i samim tim širi sferu uticaja na njenu aktivnost i mogućnost sudjelovanja u različitim reakcijama tijela. Posebno mnogo sinapsi ima u višim dijelovima nervnog sistema i to upravo u neuronima sa najsloženijim funkcijama.
U strukturi sinapse postoje tri elementa (slika 1):
- presinaptička membrana, nastao zadebljanjem membrane terminalne grane aksona;
- sinaptički rascjep između neurona;
- postsinaptička membrana zadebljanje susedne površine sledećeg neurona.
Rice. 1. Sinapsi dijagram
Pre. presinaptička membrana,
Brzo. postsinaptička membrana,
C sinoptički mjehurići,
sinoptički jaz,
M mitohondrije,
Ah acetilholin
P receptori i pore (pore)
dendrit (D) sljedeći
neuron.
Strelica jednostrano provođenje pobude.
U većini slučajeva, prijenos utjecaja s jednog neurona na drugi se provodi kemijski. U presinaptičkom dijelu kontakta postoje sinaptičkih vezikula , koji sadrže posebne supstance posrednici ili posrednici. Mogu biti acetilholin (u nekim ćelijama kičmene moždine, u vegetativnim čvorovima), norepinefrin (u završecima simpatičkih nervnih vlakana, u hipotalamusu), neki amino kiseline itd. Nervni impulsi koji pristižu na završetke aksona uzrokuju pražnjenje sinaptičkih vezikula i oslobađanje odašiljača u sinaptičku pukotinu.
Na osnovu prirode efekta na naredne nervne ćelije razlikuju se ekscitatorne i inhibitorne sinapse.
U ekscitatornim sinapsama medijatori (na primjer, acetilkolin) vezuju se za specifične makromolekule sinaptičke membrane i uzrokuju njenu depolarizaciju. U ovom slučaju, mala i kratkotrajna (oko 1 ms) fluktuacija membranskog potencijala prema depolarizacija ili ekscitatorni postsinaptički potencijal(EPSP). Da bi se neuron uzbudio, EPSP mora dostići nivo praga. Za to, depolarizacijski pomak membranskog potencijala mora biti najmanje 10 mV. Djelovanje medijatora je vrlo kratkotrajno (1-2 ms), nakon čega se dijeli na neefikasne komponente (npr. enzim kolinesteraza razlaže acetilholin u holin i sirćetna kiselina ) mulj i ponovo se apsorbira presinaptičkim terminalima (na primjer, norepinefrin).
IN inhibitorne sinapse sadržano inhibitorni medijatori (Na primjer, gama-aminobutirna kiselina ). Njihovo djelovanje na postsinaptičku membranu uzrokuje povećanje oslobađanja kalijevih jona iz stanice i povećanje polarizacije membrane. U ovom slučaju, kratkotrajna fluktuacija membranskog potencijala prema hiperpolarizacija inhibicijski postsinaptički potencijal(TPSP). Kao rezultat toga, nervna ćelija postaje inhibirana. Teže ju je uzbuditi nego u prvobitnom stanju. Ovo će zahtijevati jaču stimulaciju da bi se postigao kritični nivo depolarizacije.
Pojava impulsnog odgovora neurona
Na membrani tijela i dendritima nervnih ćelija nalaze se i ekscitatorne i inhibitorne sinapse . U određenim trenucima, neki od njih mogu biti neaktivni, dok drugi dio aktivno djeluje na susjedna područja membrane. Ukupna promjena membranskog potencijala neurona je rezultat složena interakcija(integracija) lokalnih EPSP i IPSP svih brojnih aktiviranih sinapsi. Uz istovremeni utjecaj ekscitatornih i inhibitornih sinapsa, dolazi do algebarskog sumiranja (tj. međusobnog oduzimanja) njihovih efekata. U ovom slučaju do ekscitacije neurona dolazi samo ako zbir ekscitatornih postsinaptičkih potencijala će biti više od iznosa inhibiranje . Ovaj višak mora biti određena granična vrijednost (oko 10 mV). Samo u tom slučaju se pojavljuje akcioni potencijal ćelije. Treba napomenuti da općenito ekscitabilnost neurona ovisi o njegovoj veličini: što je ćelija manja, to je veća njena ekscitabilnost .
Sa pojavom akcionog potencijala, proces vođenja nervnog impulsa duž aksona i prenoseći ga na sljedeći neuron ili radni organ, tj. vrši se efektorska funkcija neurona. Nervni impuls je glavno sredstvo komunikacije između neurona.
dakle, prijenos informacija u nervnom sistemu odvija se pomoću dva mehanizma: električnog (EPSP; IPSP; akcioni potencijal) i hemijski (posrednici).
Osobine aktivnosti nervnih centara
Svojstva nervnih centara su u velikoj meri povezana sa karakteristikama provođenja nervnih impulsa kroz sinapse koje povezuju različite nervne ćelije.
Osobine ekscitacije kroz nervne centre
Nervni centar naziva kolekcija nervnih ćelija neophodnih za obavljanje funkcije. Ovi centri odgovaraju odgovarajućim refleksnim reakcijama na vanjsku stimulaciju primljenu od receptora povezanih s njima. Ćelije nervnih centara reaguju i na njihovu direktnu iritaciju supstancama u krvi koja kroz njih teče (humoralni uticaji). U kompletnom organizmu postoji stroga koordinacija - koordinaciju njihovih aktivnosti.
Provođenje talasa pobuđivanja od jednog neurona do drugog kroz sinapsu događa se u većini nervnih ćelija hemijski uz pomoć medijatora, a medijator se nalazi samo u presinaptičkom dijelu sinapse i nema ga u postsinaptičkoj membrani. Zbog toga važna karakteristika provođenje ekscitacije kroz sinoptičke kontakte je jednostrano provođenje nervne utjecaje, što je moguće samo sa presinaptičke membrane na postsinaptičku i nemoguće je u suprotnom smjeru. S tim u vezi, tok nervnih impulsa u refleksnom luku ima određeni smjer od aferentnih neurona do interkalarnih neurona, a zatim do eferentnih motornih neurona ili autonomnih neurona.
Velika važnost u aktivnosti nervnog sistema ima još jednu osobinu provođenja ekscitacije kroz sinapse sporo provođenje. Vrijeme utrošeno na procese koji se odvijaju od trenutka kada se nervni impuls približi presinaptičkoj membrani do pojave potencijala u postsinaptičkoj membrani naziva se sinaptičko kašnjenje. U većini centralnih neurona iznosi oko 0,3 ms. Nakon toga je potrebno više vremena za razvoj ekscitatornog postsinaptičkog potencijala (EPSP) i akcionog potencijala. Cijeli proces prijenosa nervnog impulsa (od akcionog potencijala jedne ćelije do akcionog potencijala sljedeće ćelije) kroz jednu sinapsu traje otprilike 1,5 ms. Uz umor, hlađenje i niz drugih utjecaja, trajanje sinaptičkog kašnjenja se povećava. Ako neka reakcija zahtijeva učešće veliki broj neurona (mnoge stotine, pa čak i hiljade), onda ukupna vrijednost kašnjenja u provođenju kroz nervne centre može biti desetinke sekunde, pa čak i cijele sekunde.
Tokom refleksne aktivnosti ukupno vrijeme od trenutka primjene vanjske iritacije do pojave odgovora tijela tzv skriveno ili latentno vrijeme refleksa određeno uglavnom trajanjem provođenja kroz sinapse. Veličina latentnog vremena refleksa je važna indikator funkcionalno stanje nervnih centara . Mjerenje latentnog vremena jednostavne motoričke reakcije osobe na vanjski signal se široko koristi u praksi za procjenu funkcionalnog stanja centralnog nervnog sistema (slika 2).
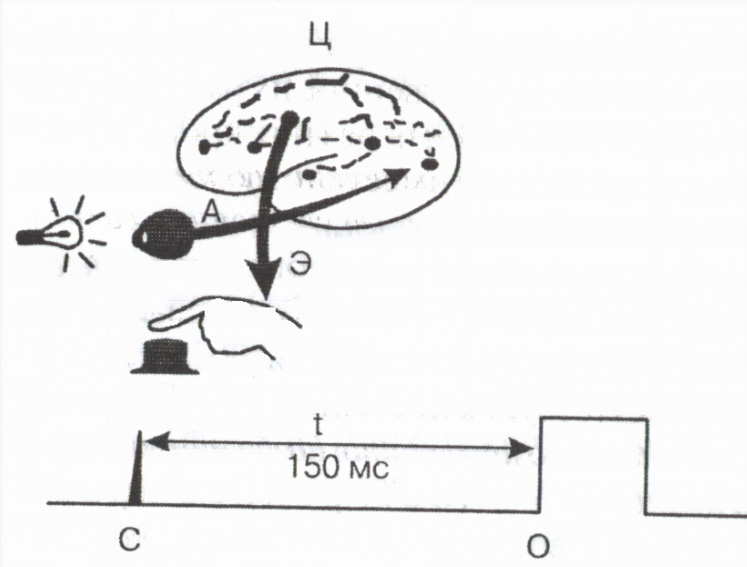
Rice. 2. Šema za mjerenje vremena motoričke reakcije
Aferent
E eferentni i C centralni putevi; C - oznaka svjetlosnog signala,
Oznaka pritiska na dugme,
t 150ms vrijeme reakcije.
Sumiranje pobude
Kao odgovor na jedan aferentni val koji putuje od receptora do neurona, mala količina transmitera se oslobađa u presinaptičkom dijelu sinapse. U ovom slučaju, EPSP se obično javlja u postsinaptičkoj membrani neurona, mala lokalna depolarizacija. Da bi ukupna vrijednost EPSP-a na cijeloj neuronskoj membrani dostigla prag za nastanak akcionog potencijala, potrebno je sumiranje mnogih podpragovnih EPSP-a na ćelijskoj membrani. Samo kao rezultat takvog zbrajanja ekscitacije nastaje odgovor neurona. Pravi se razlika između prostornog i vremenskog zbrajanja.
Prostorna sumacija uočeno u slučaju istovremenog prijema više impulsa u isti neuron duž različitih presinaptičkih vlakana. Istovremena ekscitacija sinapsi u različitim dijelovima neuronske membrane povećava amplitudu ukupnog EPSP do granične vrijednosti. Kao rezultat, javlja se impuls odgovora iz neurona i javlja se refleksna reakcija. Na primjer, dobivanje odgovora od motoričke ćelije kičmene moždine obično zahtijeva istovremenu aktivaciju 50-100 aferentnih vlakana iz odgovarajućih perifernih receptora.
Sumiranje vremena nastaje kada se isti aferentni put aktivira nizom uzastopnih podražaja. Ako su intervali između dolaznih impulsa dovoljno kratki, EPSP neurona iz prethodnih podražaja nema vremena da se raspadne, tada se naknadni EPSP-ovi superponiraju jedan na drugi sve dok depolarizacija neuronske membrane ne dostigne kritični nivo za nastanak akcije potencijal. Na ovaj način čak blage iritacije nakon nekog vremena mogu izazvati reakcije tijela (na primjer, kihanje i kašljanje kao odgovor na blage iritacije sluzokože respiratornog trakta).
Transformacija i asimilacija ritma
Priroda odgovornog pražnjenja neurona ovisi ne samo o svojstvima stimulusa, već i o funkcionalnom stanju samog neurona(njegov membranski naboj, ekscitabilnost, labilnost). Nervne ćelije imaju svojstvo da menjaju frekvenciju prenesenih impulsa, tj. svojstvo transformacije ritma.
Kada je neuron jako ekscitabilan (na primjer, nakon uzimanja kofeina), povećana brzina impulsa (animacija ritma) , a kod niske razdražljivosti (na primjer, kod umora). usporavanje ritma , budući da se nekoliko dolaznih impulsa mora sabrati da bi se konačno dostigao prag za nastanak akcionog potencijala. Ove promjene u učestalosti impulsa mogu ojačati ili oslabiti reakcije tijela na vanjske podražaje.
Uz ritmičku stimulaciju, aktivnost neurona se može podesiti na ritam dolaznih impulsa, tj. fenomen usvajanja ritma(Ukhtomsky A. A., 1928). Razvoj usvajanja ritma osigurava podešavanje aktivnosti mnogih nervnih centara pri kontroli složenih motoričkih radnji, to je posebno važno za održavanje tempa cikličkih vježbi.
Procesi praćenja
Nakon završetka stimulusa, aktivno stanje nervne ćelije ili nervnog centra obično se nastavlja neko vreme. Trajanje procesa u tragovima varira: kratko u leđnoj moždini (nekoliko sekundi ili minuta), mnogo duže u centrima mozga (desetine minuta, sati ili čak dani) i veoma dugo u moždanoj kori (do nekoliko decenija) .
Impulsi koji kruže kroz zatvorena kola neurona mogu održati jasno i kratkotrajno stanje ekscitacije u nervnom centru. Dugotrajni skriveni tragovi su mnogo složenije prirode. Pretpostavlja se da je dugotrajno očuvanje u nervnoj ćeliji tragova sa svim karakteristična svojstva stimulus se zasniva na promjeni strukture proteina koji čine ćeliju i na restrukturiranju sinaptičkih kontakata.
Posledice kratkog pulsa (u trajanju do 1 sata) su osnova tzv kratkoročno pamćenje , a dugotrajni tragovi povezani sa strukturnim i biohemijskim preuređenjima u ćelijama su osnova formiranja dugotrajno pamćenje.
Koordinacija aktivnosti centralnog nervnog sistema
Procesi koordinacije aktivnosti centralnog nervnog sistema zasnivaju se na koordinaciji dva nervna procesa: ekscitacije i inhibicije. Inhibicija je aktivan neuronski proces , koji sprečava ili potiskuje uzbuđenje.
Značaj procesa inhibicije u centralnom nervnom sistemu
Fenomen inhibicije u nervnim centrima prvi je otkrio I.M. Sechenov 1862. godine. O značaju ovog procesa on je govorio u knjizi "Refleksi mozga" (1863).
Potapanjem žabljeg kraka u kiselinu i istovremeno iritiranjem nekih dijelova mozga (npr. nanošenjem kristala kuhinjska so u predjelu diencefalona), I.M. Sechenov je primijetio oštro kašnjenje, pa čak i potpuno odsustvo "kiselinskog" refleksa kičmene moždine (povlačenje šape). Iz ovoga je zaključio da neki nervni centri mogu značajno promijeniti refleksnu aktivnost u drugim centrima, a posebno, gornji nervni centri mogu inhibirati aktivnost nižih. Opisano iskustvo ušlo je u istoriju fiziologije pod imenom Sechenov kočenje .
Inhibicijski procesi su neophodna komponenta koordinacije nervne aktivnosti. prvo, proces inhibicije ograničava širenje ekscitacije na susjedne nervne centre, što doprinosi njegovoj koncentraciji u potrebnim područjima nervnog sistema. Drugo, nastaje u nekim nervnim centrima paralelno sa ekscitacijom drugih nervnih centara, proces inhibicije pri tome isključuje nepotrebne aktivnosti ovog trenutka organi . Treće, razvoj inhibicije u nervnim centrima štiti ih od prekomjernog preopterećenja tokom rada, odnosno igra zaštitnu ulogu.
Postsinaptička i presinaptička inhibicija
Proces kočenja , za razliku od uzbuđenja, ne može se širiti duž nervnog vlakna ovo je uvijek lokalni proces u području sinaptičkih kontakata. Na osnovu mjesta nastanka razlikuju se presinaptička i postsinaptička inhibicija.
Postsinaptička inhibicija ovo su inhibitorni efekti koji se javljaju u postsinaptičkoj membrani. Najčešće je ova vrsta inhibicije povezana s prisustvom u centralnom nervnom sistemu posebnog inhibitorni neuroni. Oni su posebna vrsta interneurona u kojima terminali aksona oslobađaju inhibitorni transmiter. Jedan od ovih posrednika je gama-aminobutirna kiselina (GAM K).
Nervni impulsi koji se približavaju inhibitornim neuronima izazivaju isti proces ekscitacije u njima kao i u drugim nervnim ćelijama. Kao odgovor, normalni akcioni potencijal se širi duž aksona inhibitorne ćelije. Međutim, za razliku od drugih neurona, završeci aksona ne oslobađaju ekscitatorni, već inhibitorni transmiter. Kao rezultat toga, inhibitorne stanice inhibiraju one neurone na kojima završavaju njihovi aksoni.
Specijalni inhibitorni neuroni uključuju Renshawove ćelije u kičmenoj moždini, Purkinje ćelije u malom mozgu, košaraste ćelije u diencefalonu, itd. Na primjer, inhibitorne ćelije su od velike važnosti u regulaciji aktivnosti mišića antagonista: što dovodi do opuštanja mišića antagonista, one na taj način olakšavaju istovremenu kontrakciju mišića agonista (slika 3).
Renshaw ćelije učestvuju u regulaciji nivoa aktivnosti pojedinih motornih neurona kičmene moždine. Kada je motorni neuron uzbuđen, impulsi putuju duž njegovog aksona do mišićnih vlakana i istovremeno duž kolaterala aksona do Renshaw inhibitorne ćelije. Aksoni potonjeg se "vraćaju" na isti neuron, uzrokujući njegovu inhibiciju. Što više ekscitatornih impulsa motorni neuron šalje periferiji (a samim tim i inhibitornoj ćeliji), to je jači povratno kočenje(vrsta postsinaptičke inhibicije). Takve zatvoreni sistem djeluje kao mehanizam za samoregulaciju neurona, štiteći ga od pretjerane aktivnosti.
Purkinje ćelije malog mozga svojim inhibitornim dejstvom na ćelije subkortikalnih jezgara i matične strukture učestvuju u regulaciji mišićnog tonusa.
Basket ćelije u diencefalonu su poput kapija koje dozvoljavaju ili ne dopuštaju da impulsi odlaze do moždane kore iz različitih dijelova tijela.
Presinaptička inhibicija javlja se prije sinaptičkog kontakta u presinadičkoj regiji. Kraj aksona inhibitorne nervne ćelije formira sinapsu na kraju aksona ekscitatorne nervne ćelije, izazivajući preterano jaku depolarizaciju membrane ovog aksona, koja inhibira akcione potencijale koji ovde prolaze i na taj način blokira prenos uzbuđenje. Ova vrsta inhibicije ograničava protok aferentnih impulsa do nervnih centara, isključujući uticaje koji su strani glavni aktivnosti.
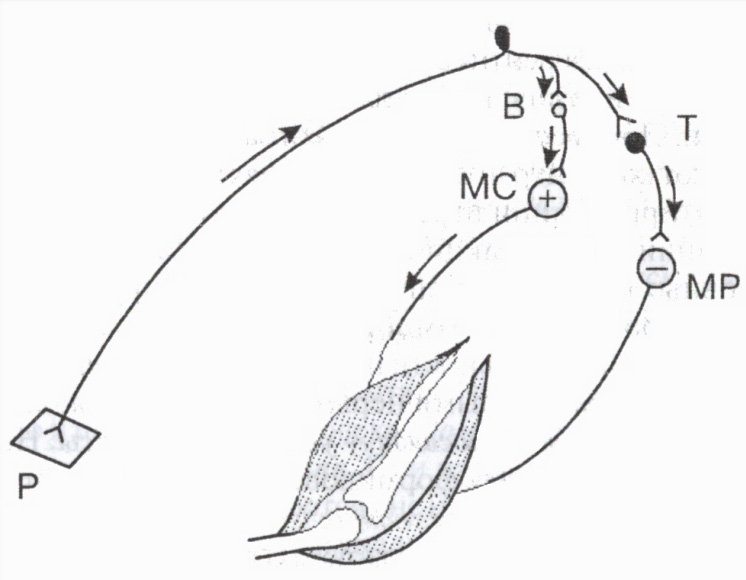
Rice. 3. Učešće inhibitorne ćelije u regulaciji mišića antagonista
B i T su ekscitatorni i inhibitorni neuroni. Ekscitacija (+) motornog neurona mišića fleksora (MS) i inhibicija (-) motornog neurona mišića ekstenzora (MR). P kožni receptor.
Fenomen zračenja i koncentracije
Kada je jedan receptor stimulisan, ekscitacija se u principu može širiti u centralnom nervnom sistemu u bilo kom pravcu i na bilo koju nervnu ćeliju. To se događa zbog brojnih međusobnih veza neurona jednog refleksni luk sa neuronima drugih refleksnih lukova. Širenje procesa ekscitacije na druge nervne centre naziva se fenomen zračenje.
Što je jača aferentna stimulacija i što je veća ekscitabilnost okolnih neurona, to više neurona pokriva proces zračenja. Procesi inhibicije ograničavaju zračenje i doprinose koncentraciji ekscitacije na početnoj tački centralnog nervnog sistema.
Proces zračenja igra važnu ulogu pozitivnu ulogu tokom formiranja novih reakcija tijela (indikativne reakcije, uvjetni refleksi). Što je više različitih nervnih centara aktivirano, lakše je među njima izabrati centre koji su najpotrebniji za naredne aktivnosti. Zahvaljujući zračenju ekscitacija između različitih nervnih centara novi funkcionalni odnosi nastaju uslovni refleksi . Na osnovu toga moguće je, na primjer, formirati nove motoričke sposobnosti.
Istovremeno, može imati i zračenje ekscitacije negativan uticaj na stanje i ponašanje tijela, narušavajući suptilne odnose između uzbuđenih i inhibiranih nervnih centara i uzrokujući poremećena koordinacija pokreta .
Dominantno
Istražujući karakteristike međucentralnih odnosa, A. A. Ukhtomsky je otkrio da ako se u tijelu životinje provodi složena refleksna reakcija, na primjer, ponovljeni činovi gutanja, tada električna stimulacija motoričkih centara ne samo da prestaje da uzrokuje pomicanje udova u ovom trenutku. momenta, ali i pojačava tok započete lančane reakcije, gutanja, za koje se pokazalo da je dominantno.
Takav dominantni fokus ekscitacije u centralnom nervnom sistemu, koji određuje trenutnu aktivnost tela, označio je A. A. Ukhtomsky (1.923) pojmom dominantan.
Dominantni fokus može se pojaviti kada povišen nivo ekscitabilnost nervnih ćelija, koju stvaraju različiti humoralni i nervni uticaji. On potiskuje aktivnost drugih centara, vršeći pridruženu inhibiciju.
Objedinjavanje velikog broja neurona u jedan dominantan sistem se dešava međusobnim usklađivanjem sa opštim tempom aktivnosti, odnosno asimilacijom ritma. Neke nervne ćelije smanjuju svoju višu stopu aktivnosti, dok druge povećavaju svoju nisku stopu do određenog prosječnog, optimalnog ritma. Dominantna može ostati dugo vremena u skrivenom stanju u tragovima (potencijalna dominanta). Kada se obnovi prethodno stanje ili prethodna vanjska situacija, dominanta može ponovo nastati (ažuriranje dominante). Na primjer, u predstartnom stanju se aktiviraju svi oni nervni centri koji su bili dio sistema rada tokom prethodnog treninga, te se shodno tome poboljšavaju funkcije vezane za rad. Mentalno izvršenje fizičke vežbe ili reprezentacija pokreta takođe reproducira radnu dominantu, koja daje trenažni efekat predstavljanja pokreta i osnova je takozvanog ideomotornog treninga. Uz potpunu relaksaciju (na primjer, s autogeni trening) sportisti nastoje eliminirati radne dominante, što ubrzava procese oporavka.
Kao faktor ponašanja, dominantno se povezuje sa najvišim nervna aktivnost i ljudska psihologija. Dominantna je fiziološku osnovučin pažnje. U prisustvu dominantne, mnogi uticaji spoljašnjeg okruženja ostaju van naše pažnje, ali oni koji nas posebno zanimaju se intenzivnije hvataju i analiziraju. Dakle, dominanta je snažan faktor u odabiru biološki i društveno najznačajnijih stimulansa.
Funkcije kičmene moždine i subkortikalnih dijelova mozga
U centralnom nervnom sistemu postoje drevniji segmentalni i evolutivno mlađi suprasegmentalni delova nervnog sistema. Segmentni dijelovi uključuju kičmenu moždinu, duguljastu moždinu i srednji mozak, čiji dijelovi reguliraju funkcije pojedinih dijelova tijela koji leže na istom nivou. Suprasegmentalni odsjeci - diencefalon, mali mozak i moždana kora nemaju direktne veze sa organima tijela, već kontroliraju njihovu aktivnost preko donjih segmentnih odjeljaka.
Kičmena moždina
Kičmena moždina je najniži i najstariji dio centralnog nervnog sistema . Siva tvar ljudske kičmene moždine sadrži oko 13,5 miliona nervnih ćelija. Od njih većina (97%) su srednje ćelije (interneuroni ili interneuroni) , koji obezbeđuju složene procese koordinacije unutar kičmene moždine. Među motornim neuronima kičmene moždine postoje veliki alfa motornih neurona i male gama motornih neurona. Najdeblja i najbrže provodna vlakna motornih nerava odlaze od alfa motornih neurona, izazivajući kontrakcije skeletni mišićna vlakna. Tanka vlakna gama motornih neurona ne uzrokuju kontrakciju mišića. Približavaju se proprioreceptorima mišićna vretena i regulišu njihovu osetljivost.
Refleksi kičmene moždine mogu se podijeliti na motor izvode alfa motorni neuroni prednjih rogova, i vegetativno izvode aferentne ćelije bočnih rogova.
Motorni neuroni kičmene moždine inerviraju sve skeletne mišiće (osim mišića lica) . Kičmena moždina izvodi elementarnih motoričkih refleksa fleksija i ekstenzija, ritmična, iskoračna, koja nastaje zbog iritacije kože ili proprioceptora mišića i tetiva, a također šalje stalne impulse mišićima, podržavajući mišićni tonus. Posebni motorni neuroni inerviraju respiratorne mišiće - interkostalne mišiće i dijafragmu, te obezbjeđuju pokreti disanja. Autonomni neuroni inerviraju sve unutrašnje organe (srce, krvne sudove, znojne žlezde, žlezde unutrašnja sekrecija, probavni trakt, genitourinarni sistem).
Funkcija provodnika Kičmena moždina je povezana s prijenosom toka informacija primljenih od periferije do gornjih dijelova nervnog sistema i sa provođenjem impulsa koji dolaze od mozga do kičmene moždine.
Iza poslednjih godina razvijen posebne tehnike za proučavanje aktivnosti kičmene moždine u zdrava osoba. Na primjer, funkcionalno stanje alfa motornih neurona procjenjuje se promjenama potencijala odgovora mišića tokom periferne stimulacije – tzv. H-refleks (Hoffmannov refleks) potkoljenični mišić kada je iritiran tibijalni nerv i po T-refleks (iz tetive tetive) mišića soleusa sa iritacijom Ahilove tetive. Razvijene su metode za snimanje (sa netaknutih površina tijela) potencijala koji prolaze kroz kičmenu moždinu u mozak.
Medulla oblongata i pons
Produžena moždina i most (zajedno zadnji mozak) dio su moždanog stabla. Postoji velika grupa kranijalnih nerava (od V do XII para) koji inerviraju kožu, sluzokožu, mišiće glave i niz unutrašnje organe(srce, pluća, jetra). Evo ih centri mnogih probavnih refleksa žvakanje, gutanje, pokreti želuca i dijela crijeva, lučenje probavnih sokova, kao i centri nekih zaštitnih refleksa (kihanje, kašalj, treptanje, suzenje, povraćanje) i centri metabolizma vode, soli i šećera . Na dnu IV ventrikula u produženoj moždini nalazi se vitalni respiratorni centar koji se sastoji od centara za udisaj i izdisaj. Sastoji se od malih ćelija koje šalju impulse respiratornim mišićima kroz motorne neurone kičmene moždine.
Nalazi se u neposrednoj blizini kardiovaskularni centar . Njegove velike ćelije regulišu rad srca i lumen krvnih sudova. Preplitanje ćelija respiratornog i kardiovaskularnog centra osigurava njihovu blisku interakciju.
Oblongata medulla igra važnu ulogu u provođenju motoričkih radnji i regulaciji tonusa skeletnih mišića, povećanje tonusa mišića ekstenzora . On posebno učestvuje u implementaciji refleksi posturalnog prilagođavanja (cervikalni, lavirint). Uzlazni ascenderi prolaze kroz produženu moždinu putevi slušne, vestibularne, proprioceptivne i taktilne osjetljivosti .
Srednji mozak
Srednji mozak se sastoji od kvadrigeminusa, crne supstance i crvenih jezgara. U prednjim tuberkulima kvadrigeminusa postoje vizuelni subkortikalni centri , i pozadi auditivni . Srednji mozak je uključen u regulacija pokreta očiju , obavlja pupilarni refleks (proširenje zenica u mraku i suženje na svetlu).
Kvadrigeminali izvode brojne reakcije, koje su komponente orijentacionog refleksa . Kao odgovor na iznenadnu iritaciju, glava i oči se okreću prema podražaju, a kod životinja se naćule uši. Ovaj refleks (prema I.P. Pavlovu, refleks "Šta je ovo?") je neophodan da se tijelo pripremi za pravovremenu reakciju na svaki novi udar.
Crna supstanca srednji mozak je povezan sa refleksima žvakanja i gutanja, uključen je u regulacija mišićnog tonusa (posebno kada se radi malim pokretima prstima) i u organizovanju prijateljskih motoričkih reakcija.
Crveno jezgro srednji mozak obavlja motoričke funkcije reguliše tonus skeletnih mišića , što uzrokuje povećan tonus mišića pregibača. Imajući značajan uticaj na tonus skeletnih mišića, srednji mozak učestvuje u brojnim instalacioni refleksi za održavanje držanja (ispravljanje postavljanja tela sa krunom nagore, itd.).
Diencephalon
Diencefalon uključuje talamus (vizualni talamus) i hipotalamus (subtalamus).
Kroz thalamus prolaze kroz sve aferentne puteve (osim olfaktornih), koji se šalju u odgovarajuća perceptivna područja korteksa (auditivna, vizuelna, itd.). Jezgra talamusa se dijele na specifične i nespecifične . Specifični uključuju komutacione (relejne) jezgre i asocijativne . Aferentni uticaji sa svih tjelesnih receptora se prenose preko preklopnih jezgara talamusa. Asocijativna jezgra primaju impulse od preklopnih jezgara i osiguravaju njihovu interakciju. Pored ovih jezgara, talamus sadrži nespecifična jezgra koja imaju i aktivirajuće i inhibitorno djelovanje na mala područja korteksa.
Zahvaljujući svojim širokim vezama, talamus igra vitalnu ulogu u funkcioniranju tijela. Impulsi koji dolaze iz talamusa u korteks mijenjaju stanje kortikalnih neurona i regulišu ritam kortikalne aktivnosti . Uz direktno učešće talamusa javlja se formiranje uslovnih refleksa i razvoj motoričkih sposobnosti, formiranje emocija osobu, njene izraze lica. Talamus posebno igra veliku ulogu u nastanku osjeta senzacije bola . Njegove aktivnosti su povezane regulacija bioritma u životu osobe (svakodnevni, sezonski, itd.).
Hipotalamus je najviši subkortikalni centar regulacije vegetativne funkcije , navodi budnost i san . Ovdje se nalaze vegetativni centri, regulisanje metabolizma i telo, obezbeđujući održavanje stalne tjelesne temperature (kod toplokrvnih životinja) i normalan nivo krvni pritisak , podržavajući bilans vode regulisanje osećaj gladi i sitosti . Iritacija stražnjih jezgara hipotalamusa uzrokuje povećanje simpatičkih utjecaja, a prednjih - parasimpatičkih.
Zbog povezanosti hipotalamusa i hipofize ( hipotalamo-hipofiznog sistema) kontroliše se aktivnost endokrinih žlijezda. Autonomne i hormonalne reakcije, koje regulira hipotalamus, komponente su ljudskih emocionalnih i motoričkih reakcija.
Nespecifični moždani sistem
Nespecifičan sistem zauzima srednji dio moždanog stabla. Ne uključuje analizu bilo koje specifične osjetljivosti ili izvođenje specifičnih refleksnih reakcija. Impulsi u ovaj sistem ulaze kroz bočne grane sa svih specifičnih puteva, što rezultira njihovom ekstenzivnom interakcijom. Za nespecifičan sistem Karakteristična je lokacija neurona u obliku difuzne mreže, obilje i raznolikost njihovih procesa. U tom smislu je dobio naziv retikularna formacija ili retikularna formacija.
Razlikovati dve vrste uticaja nespecifičan sistem na rad drugih nervnih centara aktivirajuće i inhibitorno . Obje vrste ovih utjecaja mogu biti uzlazne (do gornjih centara) i silazne (do temeljnih centara). Oni služe za regulacija funkcionalnog stanja mozga, nivoa budnosti i regulacija posturalno-toničnih i faznih reakcija skeletnih mišića.
Mali mozak
Mali mozak je suprasegmentalna formacija koja nema direktne veze sa izvršnim aparatom. Mali mozak se sastoji od neuparene formacije - vermisa i uparenih hemisfera.
Glavni neuroni malog korteksa su brojni Purkinje ćelije. Zahvaljujući ekstenzivnim vezama (svaka ćelija ima do 200.000 sinapsi), sadrže integraciju širokog spektra senzornih uticaja , prvenstveno proprioceptivni, taktilni i vestibularni. Reprezentacija različitih perifernih receptora u korteksu malog mozga ima somatotopska organizacija (grč. somatos tijelo, topos mjesto), tj. odražava redoslijed njihovog položaja u ljudskom tijelu. Pored toga, ovaj redosled rasporeda odgovara istom redosledu rasporeda predstavljanja delova tela u kori velikog mozga, što olakšava razmenu informacija između korteksa i malog mozga i obezbeđuje njihovu zajedničku aktivnost u kontroli ljudskog ponašanja. Ispravna geometrijska organizacija neurona malog mozga određuje njegovu važnost u računanju vremena i jasnom održavanju tempa cikličnih pokreta.
Glavna funkcija malog mozga je regulacija posturalnih reakcija i koordinacija motoričke aktivnosti.(Orbeli L.A., 1926).
By anatomske karakteristike(veze korteksa malog mozga sa njegovim jezgrima) i funkcionalnog značaja, mali mozak se dijeli na tri uzdužne zone:
- unutrašnji ili medijalni korteks vermisa, čija je funkcija regulacija tonusa skeletnih mišića, održavanje držanja tijela i ravnoteže;
- srednji srednji dio cerebralni korteks mali mozak, čija je funkcija da koordinira posturalne reakcije s pokretima i ispravlja greške;
- lateralni ili lateralni korteks hemisfera malog mozga, koji zajedno sa diencephalon a cerebralni korteks je uključen u programiranje brzih balističkih pokreta (bacanja, udarci, skokovi itd.).
Bazalni gangliji
Bazalni gangliji uključuju striatum, koji se sastoji od kaudatnog jezgra i putamena, i blijedi nukleus, a sada se također smatraju amigdala(vezano za vegetativni centri limbički sistem) i supstancija nigra srednjeg mozga.
Aferentni utjecaji dolaze do bazalnih ganglija od tjelesnih receptora preko talamusa i iz svih područja kore velikog mozga. Gotovo isključivo ulaze u striatum. Eferentni uticaji iz njega se usmeravaju na blijedo jezgro i dalje ka stabnim centrima ekstrapiramidnog sistema, kao i kroz talamus nazad u korteks.
Bazalni gangliji su uključeni u formiranje uslovnih refleksa i implementaciju kompleksa bezuslovnih refleksa(odbrambeni, za proizvodnju hrane, itd.). Obezbeđuju neophodan položaj tela tokom fizičkog rada, kao i protok automatskih ritmičkih pokreta (drevni automatizmi).
Palidus jezgra obavlja glavnu motoričku funkciju, a striatum regulira njegovu aktivnost. Trenutno je otkrivena važnost kaudatnog jezgra u kontroli složenih mentalnih procesa – pažnje, pamćenja i detekcije grešaka.
Sve funkcije tijela mogu se podijeliti na somatske, ili životinja(životinje) povezane s percepcijom eksterne informacije i mišićna aktivnost, i vegetativna (biljna) povezana sa aktivnostima unutrašnjih organa, procesima disanja, cirkulacije krvi, probave, izlučivanja, metabolizma, rasta i razmnožavanja.
Funkcionalna organizacija autonomnog nervnog sistema
Autonomni nervni sistem naziva se skup eferentnih nervnih ćelija kičmene moždine i mozga, kao i ćelije posebnih čvorova (ganglija) koji inerviraju unutrašnje organe. Stimulacija različitih tjelesnih receptora može uzrokovati promjene i somatskih i autonomnih funkcija, jer su aferentni i centralni dijelovi ovih refleksnih lukova uobičajeni. Razlikuju se samo po svojim eferentnim dijelovima. Karakteristična karakteristika eferentni putevi uključeni u refleksne lukove autonomnih refleksa su njihova struktura od dva neurona(jedan neuron se nalazi u centralnom nervnom sistemu, drugi u ganglijama ili u inerviranom organu).
Vegetativno nervni sistem podijeljen u dva dijela: simpatikus i parasimpatikus (Sl. 4).
Eferentni putevi simpatičkog nervnog sistema počinju u grudima i lumbalne regije kičmene moždine od neurona njenih bočnih rogova. Prijenos ekscitacije s prenodalnih simpatičkih vlakana na postnodalna vlakna događa se uz sudjelovanje posrednika acetilholin , te od postnodalnih vlakana do inerviranih organa uz učešće posrednika norepinefrin . Izuzetak su vlakna koja inerviraju znojne žlijezde i proširuju žile skeletnih mišića, gdje se ekscitacija prenosi acetilkolinom.
Eferentni putevi parasimpatičkog nervnog sistema počinju u mozgu od nekih jezgri srednjeg i oblongata medulla, te u kičmenu moždinu od neurona sakralni region. Provođenje ekscitacije na sinapsama parasimpatičkog puta odvija se uz sudjelovanje medijatora acetilholin . Drugi eferentni neuron nalazi se u ili blizu inerviranog organa.
Najviši regulator autonomnih funkcija je hipotalamus , koji djeluje u sprezi sa retikularnom formacijom i limbičkim sistemom pod kontrolom moždane kore. Osim toga, neuroni smješteni u samim organima ili u simpatički čvorovi, mogu izvoditi sopstvene refleksne reakcije bez učešća centralnog nervnog sistema "periferni refleksi" .
Rice. 4. Autonomni nervni sistem
Lijevo izlazno područje vlakana: parasimpatičko (crno)
i simpatički (zasjenjeni) sistemi.
Desno je struktura eferentnog dijela refleksnog luka autonomnih refleksa. Na lijevoj strani je dijagram sredine, duguljaste moždine i kičmene moždine.
Arapski brojevi su brojevi torakalnih segmenata, rimski brojevi lumbalnih segmenata.
Funkcije simpatičkog nervnog sistema
Uz sudjelovanje simpatičkog nervnog sistema, u tijelu se javljaju mnogi važni refleksi koji imaju za cilj osiguranje njegovog aktivnog stanja, uključujući njegovu motoričku aktivnost. To uključuje reflekse proširenje bronha, ubrzan rad srca i intenziviranje , širenje krvnih sudova srca i pluća uz istovremeno sužavanje krvnih sudova kože i organa trbušne duplje (osiguravanje preraspodjele krvi ), oslobađanje uskladištene krvi iz jetre i slezene, razlaganje glikogena u glukozu u jetri ( mobilizacija izvora energije ugljikohidrata ), povećana aktivnost endokrinih i znojnih žlezda. Simpatički nervni sistem smanjuje aktivnost brojnih unutrašnjih organa: kao rezultat vazokonstrikcije u bubrezima, procesi stvaranja urina su smanjeni, sekretorna i motorička aktivnost organa inhibirana gastrointestinalnog trakta; čin mokrenja je spriječen, zidni mišić se opušta Bešika i njegovog sfinktera.
Povećanu aktivnost tijela prati simpatička refleks širenja zenice . Od velikog je značaja za motoričku aktivnost organizma trofički utjecaj simpatičkih živaca na skeletne mišiće , poboljšavaju njihov metabolizam i funkcionalno stanje, ublažavaju umor.
Simpatički nervni sistem ne samo da povećava nivo funkcionisanja organizma, već i mobilizira svoje skrivene funkcionalne rezerve , aktivira moždanu aktivnost, pojačava zaštitne reakcije ( imunološke reakcije, mehanizme barijere itd.), izaziva hormonske reakcije. Od posebnog značaja je simpatički nervni sistem tokom razvoja stresnim uslovima , u većini teški usloviživotna aktivnost. L. A. Orbeli je naglasio vitalni značaj simpatički uticaji za adaptaciju (prilagođavanje) organizma na naporan rad, na različitim uslovima spoljašnje okruženje. Ovu funkciju je on nazvao adaptivno-trofički.
Funkcije parasimpatičkog nervnog sistema
Parasimpatički nervni sistem obavlja suženje bronha, usporavanje i slabljenje srčanih kontrakcija: sužavanje krvnih sudova srca; obnavljanje energetskih resursa (sinteza glikogena u jetri i jačanje procesa probave); jačanje procesa stvaranja urina u bubrezima i obezbeđivanje čina mokrenja (kontrakcija mišića bešike i opuštanje njenog sfinktera) itd. Parasimpatički nervni sistem uglavnom obezbeđuje pokretački uticaji : suženje zjenice, bronha, aktivacija aktivnosti probavne žlezde i tako dalje.
Aktivnost parasimpatičkog odjela autonomnog nervnog sistema usmjerena je na trenutnu regulacija funkcionalnog stanja, za održavanje postojanosti unutrašnje okruženje homeostaza . Parasimpatički odjel obezbjeđuje obnavljanje različitih fizioloških pokazatelja , naglo promijenjen nakon intenzivnog rad mišića, nadopunjavanje potrošenih energetskih resursa. Posrednik parasimpatički sistem acetilholin, smanjujući osjetljivost adrenergičkih receptora na djelovanje adrenalina i norepinefrina, ima određenu antistresno dejstvo .
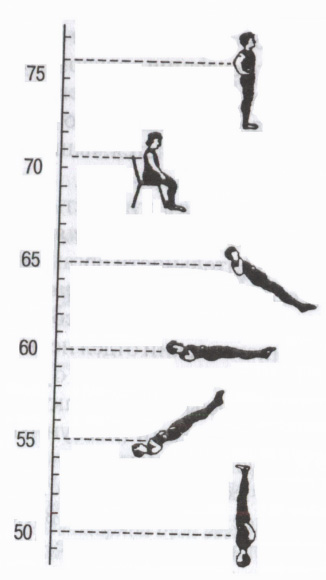
Rice. 5. Autonomni refleksi
Utjecaj položaja tijela na broj otkucaja srca (bpm). (Prema: Mogendovich M.R., 1972)
Kroz autonomne simpatičke i parasimpatičke puteve, centralni nervni sistem izvodi neke autonomne reflekse, počevši od različitih receptora spoljašnje i unutrašnje sredine: viscero-visceralni (od unutrašnjih organa do unutrašnjih organa - na primer, respiratorno-srčani refleks); dermo-visceralni (sa kože - promjene u aktivnosti unutrašnjih organa tokom iritacije aktivne tačke kože, na primjer, akupunktura, akupresura); iz receptora očne jabučice okulo-kardijalni Aschnerov refleks (smanjenje otkucaja srca pri pritisku na očne jabučice parasimpatički efekat); motorno-visceralni - na primjer, ortostatski test(povećan rad srca pri prelasku iz ležećeg u stojeći položaj simpatički efekat) itd. (Sl. 5). Koriste se za procjenu funkcionalnog stanja organizma, a posebno stanja autonomnog nervnog sistema (procjena uticaja njegovog simpatičkog ili parasimpatičkog odjela).
Limbički sistem
Ispod limbički sistem razumiju niz kortikalnih i subkortikalnih struktura čije su funkcije vezane za organizaciju motivaciono-emocionalne reakcije, pamćenje i procesi učenja .
Kortikalni dijelovi limbičkog sistema, koji predstavljaju njegov najviši dio, nalaze se na donjim i unutrašnjim površinama moždanih hemisfera (dijelovi frontalnog korteksa, cingularnog girusa ili limbičkog korteksa, hipokampusa itd.). TO subkortikalne strukture Limbički sistem uključuje hipotalamus, neka jezgra talamusa, srednji mozak i retikularnu formaciju. Između svih ovih odjela postoje bliske direktne i povratne veze, formirajući takozvani „limbički prsten“.
Limbički sistem je uključen u širok spektar manifestacija aktivnosti organizma: u regulaciji ponašanja u ishrani i piću, ciklusu spavanja i buđenja, u procesima formiranja memorijskog traga (taloženje i izvlačenje iz memorije), u razvoju agresivno-odbrambenih reakcija , osiguravajući selektivno ponašanje. Ona stvara pozitivne i negativne emocije sa svim motornim, autonomnim i hormonskim komponentama. Električna stimulacija različitih dijelova limbičkog sistema ugrađenim elektrodama (u eksperimentima na životinjama, u klinici kod liječenja pacijenata) otkrila je prisustvo centrima zadovoljstva, formiranje pozitivnih emocija, i nezadovoljstvo, formiranje negativnih emocija. Izolovana iritacija takvih tačaka u dubokim strukturama ljudskog mozga izazvala je pojavu osjećaja „bezrazorne radosti“, „besmislene melanholije“ i „nerazumnog straha“.
Funkcije kore velikog mozga
Kod viših sisara i ljudi, vodeći dio centralnog nervnog sistema je moždana kora.
Kortikalni neuroni
Korteks je sloj sive tvari debljine 2-3 mm, koji u prosjeku sadrži oko 14 milijardi nervnih ćelija. Ono što je karakteristično za njega je obilje interneuronskih veza čiji se rast nastavlja do 18. godine, a u nekim slučajevima i kasnije.
Glavni tipovi kortikalnih ćelija su piramidalni i zvezdasti neuroni. Zvjezdasti neuroni povezan s procesima percepcije iritacija i objedinjavanjem aktivnosti različitih piramidalnih neurona.
Piramidalni neuroni provode eferentnu funkciju korteksa (uglavnom kroz piramidalni trakt) i viutrikortikalne procese interakcije između neurona udaljenih jedan od drugog. Najveće piramidalne ćelije, gigantske Betzove piramide, nalaze se u prednjem središnjem girusu (motorno područje korteksa).
Funkcionalna jedinica korteksa je vertikalna kolona međusobno povezani neuroni. Velike piramidalne stanice izdužene okomito s neuronima smještenim iznad i ispod njih formiraju funkcionalne asocijacije neurona. Težina Neuroni vertikalnog stuba odgovaraju na istu aferentnu stimulaciju (od istog receptora) istom reakcijom i zajedno formiraju eferentne odgovore piramidalnih neurona. Po potrebi, vertikalni stubovi se mogu kombinovati u veće formacije, omogućavajući složene reakcije.
Funkcionalni značaj različitih kortikalnih polja
Prema strukturnim karakteristikama i funkcionalnom značaju pojedinih kortikalnih područja cela kora je podeljena u tri glavne grupe polja primarnog, sekundarnog i tercijarnog (Sl. 6).
Primarna polja povezana sa senzornim organima i organima kretanja na periferiji. Oni pružaju pojavu senzacija . To uključuje, na primjer, polje boli i mišićno-zglobne osjetljivosti u stražnjem središnjem girusu korteksa, vidno polje u okcipitalnoj regiji, slušno polje u temporalnoj regiji i motorno polje u prednjem centralnom girusu. U primarnim poljima postoje visoko specijalizovane determinantne ćelije ili detektori, selektivno reagujući samo na određene podražaje. Na primjer, u vidnom korteksu postoje detektorski neuroni koji se pobuđuju samo kada se svjetlo uključi ili isključi, osjetljivi su samo na određeni intenzitet, na određene intervale izlaganja svjetlosti, na određenu valnu dužinu itd. Kada su primarna polja korteksa uništena, dolazi do tzv. kortikalne sljepoće, kortikalne gluvoće itd.
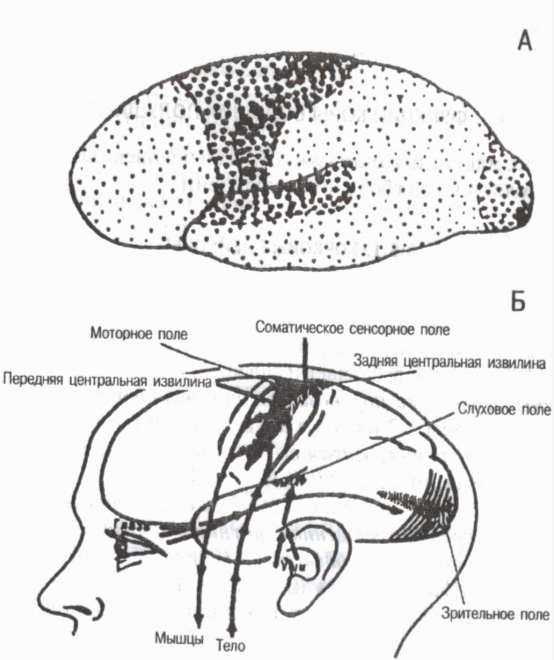
Rice. 6. Primarna, sekundarna i tercijarna polja kore velikog mozga
Na A: primarna polja velikih tačaka, srednje sekundarna polja, mala tercijarna polja (prema G. I. Polyakov 1964, A. R. Luria, 1971)
Na B: primarna (projekcijska) polja moždane kore (prema V. Penfield, L. Robert, 1964)
Sekundarna polja koji se nalaze pored primarnih. Šta se dešava u njima razumijevanje i prepoznavanje zvučnih, svjetlosnih i drugih signala, nastaju složeni oblici generalizirane percepcije . Kada su sekundarna polja oštećena, sposobnost da se vide objekti i čuju zvukovi se zadržava, ali ih osoba ne prepoznaje i ne pamti značenje.
Tercijarna polja razvijala gotovo isključivo kod ljudi. To su asocijativna područja korteksa, koja pružaju više oblike analize i sinteze i formiraju svrsishodnu aktivnost ljudskog ponašanja. Tercijarna polja se nalaze u zadnjoj polovini korteksa interparijetalnog, okcipitalnog i temporalnog regiona, a u prednjoj polovini u prednjim delovima frontalne oblasti. Njihova uloga je posebno velika u organizovanju koordinisanog rada obe hemisfere . Tercijarna polja sazrevaju kod ljudi kasnije od drugih kortikalnih polja i degradiraju ranije od ostalih tokom starenja.
Funkcija stražnjih tercijarnih polja (uglavnom donjih parijetalnih područja korteksa) je primanje, obrada i pohranjivanje informacija. Oni formiraju ideju o dijagram tijela i dijagram prostora , omogućavajući prostornu orijentaciju pokreta. Prednja tercijarna polja (frontalna područja) vrši opštu regulaciju složenih oblika ljudskog ponašanja, formiranje namjera i planova, programa dobrovoljnih pokreta i praćenje njihove realizacije . Povezan je razvoj tercijarnih polja kod ljudi sa govornom funkcijom. Razmišljanje ( unutrašnji govor) moguće je samo uz zajedničku aktivnost različitih senzornih sistema, do integracije informacija iz kojih dolazi u tercijarnim poljima. Uz urođenu nerazvijenost tercijarnih polja, osoba nije u stanju ovladati govorom (izgovara samo besmislene zvukove), pa čak ni najjednostavnijim motoričkim vještinama (ne može se oblačiti, koristiti alate itd.).
Aktivnost para i hemisferna dominacija
Kao rezultat toga, vrši se obrada informacija uparene aktivnosti obe hemisfere mozak. Međutim, po pravilu, jedna od hemisfera je vodeća dominantna . Većina ljudi sa vođenjem desna ruka(desnoruke) Dominantna je lijeva hemisfera, a podređena (subdominantna) desna hemisfera.
Lijeva hemisfera u poređenju sa desnim ima finiju neuralnu strukturu, veće bogatstvo neuronskih veza, koncentrisaniji prikaz funkcija i Bolji uslovi snabdevanje krvlju U lijevoj dominantnoj hemisferi nalazi se motorički govorni centar (Brokin centar) koji obezbjeđuje govorna aktivnost, i čulni centar govora, koji razumije riječi. Lijeva hemisfera je specijalizovana za finu senzomotornu kontrolu pokreta ruku.
Kod ljudi postoje tri oblika funkcionalna asimetrija: motoričke, senzorne i mentalne . Tipično, osoba ima dominantnu ruku, nogu, oko i uho. Međutim, problem funkcionalne asimetrije je prilično složen. Na primjer, dešnjak može imati dominantno lijevo oko ili lijevo uho, signali iz kojih su dominantni. Štaviše, u svakoj hemisferi mogu biti predstavljene funkcije ne samo suprotne, već i iste strane tijela. Kao rezultat toga, moguće je zamijeniti jednu hemisferu drugom u slučaju oštećenja, a također i stvara strukturna osnova za varijabilnu hemisfernu dominaciju u motornoj kontroli.
Mentalna asimetrija se manifestuje u obliku određenog specijalizacije hemisfere. Za leva hemisfera karakterišu analitički procesi, sekvencijalna obrada informacija, uključujući uz pomoć govora, apstraktno mišljenje, procena privremenih odnosa, anticipacija budućih događaja, uspešno rešavanje verbalnih i logičkih problema. IN desna hemisfera informacije se obrađuju holistički, sintetički (bez razlaganja na detalje), uzimajući u obzir prošlo iskustvo i bez sudjelovanja govora; prevladava supstancijalno mišljenje. Ove karakteristike omogućavaju povezivanje percepcije prostornih karakteristika i rešavanja vizuelnoprostornih problema sa desnom hemisferom. Funkcije desne hemisfere povezane su s prošlim vremenom, a lijeve hemisfere s budućnošću.
Električna aktivnost kore velikog mozga
Promjene u funkcionalnom stanju korteksa ogledaju se u njegovom snimanju električna aktivnost elektroencefalogram (EEG). Moderni elektroencefalografi pojačavaju moždane potencijale 2-3 miliona puta i omogućavaju istovremeno proučavanje EEG-a iz više tačaka korteksa, tj. sistemski procesi. EEG registracija proizveden u obliku pisanja mastilom na papiru, kao iu obliku kompletna slika na dijagramu površine mozga, odnosno moždanim mapama (metoda mapiranja) na ekranu monitora savremenih kompjuterskih sistema (slika 7).
Postoje određene frekventni opsezi , zvao EEG ritmovi (Sl. 8): u stanju relativnog mirovanja najčešće se bilježi alfa ritam (8-13 oscilacija u 1 s); u stanju aktivne pažnje - beta ritam (14 oscilacija na 1s i više); kada zaspi, neki emocionalna stanja beta ritam (4-7 oscilacija u 1s); u dubokom snu, gubitak svijesti, anestezija, delta ritam (1-3 fluktuacije u 1 s).
EEG odražava karakteristike interakcije kortikalnih neurona tokom mentalnog i fizički rad (Livanov M.N., 1972). Nedostatak dobro uspostavljene koordinacije pri obavljanju neobičnih ili teških poslova dovodi do tzv EEG desinhronizacija brza asinhrona aktivnost. Kako se motorička vještina razvija, pojave se pojavljuju u EEG-u EEG sinhronizacija povećanje međusobne povezanosti (sinhronije i u fazi) električne aktivnosti različitih područja korteksa uključenih u kontrolu pokreta. U toku cikličkog rada javljaju se spori potencijali u tempu izvedenog, imaginarnog ili nadolazećeg pokreta "označeni ritmovi"(Sologub E.B., 1973).
Osim pozadinske aktivnosti u EEG-u, odvojeno potencijali povezani sa bilo kojim događajem: evocirani potencijali, koji nastaju kao odgovor na vanjske podražaje (slušne, vizuelne, itd.); potencijali koji odražavaju moždane procese tokom pripreme, realizacije i završetka pojedinih motoričkih činova "val iščekivanja" ili uslovljeni negativni talas (Walter G., 1966), premotorni, motorni i finalni potencijali itd. ultraspore oscilacije u trajanju od nekoliko sekundi do desetina minuta (posebno tzv. “omega potencijali” itd.), koji odražavaju biohemijske procese regulacije funkcija i mentalne aktivnosti.

Rice. 7. Mapiranje mozga
Višekanalno snimanje ljudskog elektroencefalograma (EEG) na ekranu monitora i refleksije ekscitiranih (svjetle zone) i inhibiranih (tamne zone) područja korteksa.
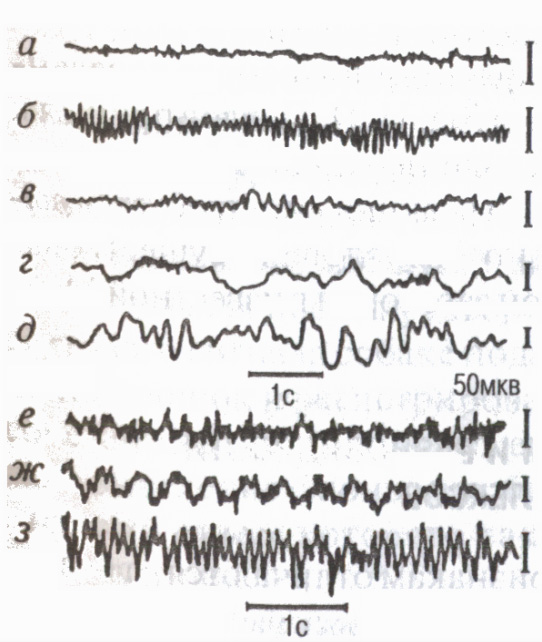
Rice. 8. EEG okcipitalnog (a-e) i motornog (f-h) područja korteksa ljudskog mozga tokom razne države i tokom rada mišića
aktivno stanje,
oči otvorene (beta ritam); b- mirovanje, zatvorene oči (alfa ritam);
c pospanost (tesha-ritam);
d zaspati; d dubok san„H
(delta ritam); e neobično ili
naporan rad asinhroni čest
aktivnost (fenomen desinhronizacije);
g ciklični rad sporih potencijala u tempu pokreta ("označeni ritmovi" EEG); h izvođenje savladanog pokreta izgleda alfa ritma.
Medicina i veterina
Piramidalni neuroni. Velike ćelije, perikarya veličine 10-130 mikrona. Ćelija ima apikalni dendrit, koji je usmjeren prema molekularnom sloju; bočni nastavci – dendriti; dugačak akson dolazi iz baze - to će biti motorno nervno vlakno (početak formiranja piramidalnog trakta).
Predavanje iz histologije br. 13
Centralni nervni sistem. Cerebralni korteks. Citoarhitektura slojeva cerebralnog korteksa. Neuronski sastav. Karakteristike piramidalnih neurona. Modul strukturne i funkcionalne jedinice kore velikog mozga. Mali mozak. Struktura i neuralni sastav kore malog mozga.
Izvor razvoja sive i bijele tvari mozga
Razvija se iz plašta ili zone plašta sive tvari neuralne cijevi
Marginalni veo bijele tvari
Komponente siva tvar mozga (neokorteks)
6 slojeva:
- Molekularno se formira uglavnom procesima ćelija ispod slojeva ispod, ali postoje pojedinačni neuroni koji se nazivaju horizontalni.
- Eksterna zrnasta prisutna u obliku zvijezde neurona i ne brojnih piramidalnih neurona.
- Vanjski piramidalni (piramidalni) najširi sloj, sadržan piramidalnih neurona.
- Unutrašnja granulacija fino u obliku zvijezde neuroni izvode intrakortikalne veze (ne ulaze u bijelu tvar).
- Unutrašnja piramidalna (ganglijska) je predstavljena velikim piramidalni neurona ili Bettsovih ćelija.
- Polimorfni oblici ćelija su raznoliki, ćelije su male, njihovi aksoni se protežu u bijelu tvar.
Zvjezdasti neuroni.Perikarije imaju okrugli ili trokutasti oblik, promjera 4-8 mikrona; akson i dendriti ovih ćelija učestvuju u intrakortikalnim vezama.
Piramidalni neuroni.Velike ćelije, perikarija veličine 10-130 mikrona. Ćelija ima apikalni dendrit, koji je usmjeren prema molekularnom sloju; bočni procesi dendriti; dugačak akson dolazi iz baze i bit će motorno nervno vlakno (početak formiranja piramidalnog trakta). Kolaterali dolaze iz aksona: idu u susjedne ćelije ili se podižu i stupaju u interakciju sa slojevima koji su iznad.
opšte karakteristike kortikalni neuroni:
Svi neuroni kore velikog mozga su multipolarni. Po funkciji, slojevi 1-4: asocijativni (interkalarni), 5-6 motorni.
Strukturna i funkcionalna jedinica mozga: modul
Modul (u korteksu velikog mozga do 3 miliona) vertikalni stub prečnika 300 mikrona, okomit na površinu korteksa i prolazi kroz celu debljinu korteksa.
U sredini modula:
- Jedno kortikokartikularno nervno vlakno je nervno vlakno unutar korteksa; dolaze ili iz piramidalnih neurona iste hemisfere (asocijativno-interkalarni) ili iz suprotne hemisfere (komisuralni).
- Dva talamokortikalna vlakna (vlakna osjetljiva na afirmaciju), koja dolaze iz thalamus opticusa i idu u 4 sloja; tada se formira eferentno nervno vlakno formiran od aksona Najbolje ćelije.
Citoarchitecturepodručja korteksa koja se međusobno razlikuju po strukturi i lokaciji stanica.
Myeloarchitecturepodručja korteksa koja se međusobno razlikuju po karakteristikama rasporeda vlakana.
Polja različita područja korteksa koja se međusobno razlikuju: cito, mijeloarhitektura i funkcionalni značaj.
Vrste kore:
- Zrnasti osjetljivi centar, u kojem su slojevi 2 i 4 dobro razvijeni; završavaju (aferentna) senzorna nervna vlakna koja dolaze iz organa mirisa, sluha i vida.
- Agranularni motorni centar, u kojem su razvijeni slojevi 3, 5 i 6.
Pouzdan morfološki ekvivalent inteligencije je broj sinapsi asocijativnih neurona (slojevi 1-4), obično 10 hiljada.
Mali mozak
Izvor razvoja zadnjeg mozga malog mozga
Funkcije: koordinira kretanje i ravnotežu
Mali mozak ima veliki broj konvolucija, u preparaciji u obliku razgranatog stabla.
Vijuge su odvojene žljebovima. Svaki girus sadrži: usku ploču bijele tvari, potpuno prekrivenu siva tvar(kora). Grupe neurona leže duboko u bijeloj tvari malog mozga.: jezgra malog mozga.
Siva tvar ili cerebelarni korteks ima tri sloja:
- Eksterna molekula je predstavljena multipolarnim asocijativnim (interkalarnim) neuronima. Postoje 2 vrste ćelija:
- Basket neuroni se nalaze u donjoj trećini molekularnog sloja, ćelija nepravilnog oblika male veličine. Akson ćelije sinapsira sa perikareonom Purkenje ćelije, gde se grana i formira pleksus u obliku korpe. Dendriti u molekularnom sloju.
- Zvjezdaste (velike i male) lokalizirane su s vanjske strane molekularnog sloja (površine korteksa).Veliki zvezdasti neuroni:akson sinapsira sa perikarijom Purkenje ćelija i učestvuje u formiranju korpe. Dendriti u molekularnom sloju.Mali zvezdasti neuroni: Akson sinapsira sa dendritima Purkenje ćelija.
- Srednji ganglijski - predstavljen jednim slojem, tijelima multipolarnih motornih neurona ili Purkinje ćelija. To su velike ćelije u obliku kruške iz perikariona (tijela) sa 2-3 dendrita koji se protežu u molekularni sloj. Akson nastaje iz tijela ćelije i prolazi kroz granularni sloj u bijelu tvar, gdje se završava na jezgri malog mozga. Aksoni Purkinjeovih ćelija su jedina eferentna (motorna) nervna vlakna koja izlaze iz korteksa malog mozga. Bliže tijelu ćelije, od aksona odlaze kolaterali, koji idu do ganglijskog sloja i dubokih dijelova molekularnog sloja, gdje sinapsiraju s tijelima i dendritima.susjedne Purkinje ćelije.
- Unutrašnje granularne sve ćelije su polarne i asocijativne sadrže dvije vrste ćelija:
1-Ćelije-Zrna male ćelije, perikarioni imaju prečnik 4-6 mikrona, dendriti: kratki 3-4 komada, usmereni ka beloj materiji, podsećaju na ptičje noge. Akson se uzdiže u molekularni sloj, gdje se dijeli u obliku slova T.
2-Velika zvezdasta: sa dugim aksonima i kratkim aksonima Golgijeve ćelije drugog tipa. Velike ćelije tela lokalizovane su neposredno ispod ganglijskog sloja. Akson sinapsira s rozetama mahovinastih vlakana prije nego što sinapsira s dendritima granularnih stanica. Dendriti se dižu u molekularni sloj.
Aferentna (osjetljiva) nervna vlakna:
- Mahovina vlakna mahovinasti terminal nervno vlakno se zove utičnica. Rozete od mahovinastih vlakana sinapse sa dendritima ćelija granula, formirajući cerebelarne glomerule.
- Vlakna koja se penjaju ili u obliku lijane dolaze iz bijele tvari, prolaze kroz granularne slojeve i sinapse ili sa perikarionom Purkinje ćelije ili sa dendritima ovih ćelija.
Aksoni granularnih ćelija sinapse sa dendritima:
- Molekularni sloj ćelije
- Purkenje ćelije
- Golgijeva ćelija tip II
Inhibicijski sistem malog mozga uključuje:
- Ćelije molekularnog sloja
- Golgijeve ćelije tipa 2
- Ekscitatorne sinapse:
- Mahovinasta vlakna
- Liana vlakna
Kao i ostali radovi koji bi vas mogli zanimati |
|||
| 73026. | Unošenje, formatiranje podataka i pisanje formula | 126 KB | |
| Cilj: Koristite naredbu Format Cells da formatirate podatke u koloni D u valutnom formatu bez decimalnih mjesta. Koristeći dugme Format Pattern, kopirajte format kolone D u E. Koristite dugmad na panelu Formatting da biste naveli procentualni format sa dve decimale u koloni F. | |||
| 73027. | Izrada i uređivanje grafikona u Excel-u | 160 KB | |
| Naučite da pravite dijagrame pomoću čarobnjaka; Naučite uređivati dijagrame. Test pitanja Koja je svrha dijagrama Koje vrste dijagrama znate Kako napraviti dijagram poseban list Kako promijeniti vrstu grafikona Kako izbrisati grafikon... | |||
| 73028. | Modeliranje sistema datoteka | 147,5 KB | |
| Korisnici daju fajlovima simbolična imena, uzimajući u obzir određena OS ograničenja. Direktorij sadrži listu datoteka uključenih u njega i uspostavlja korespondenciju između datoteka i njihovih karakteristika i atributa. | |||
| 73029. | Određivanje strukturno-faznog sastava NVM, koji zamjenjuje VNT, metodama rendgenske difrakcije i elektronske mikroskopije | 1,5 MB | |
| Važna je strukturno-fazna struktura NVM-a kako bi se VNT uskladio sa podacima rendgenske difrakcije i elektronske mikroskopije. Posmatrajte promjenu strukturnog faznog sastava NVM tokom termohemijske obrade. | |||
| 73033. | Prilagođeni tipovi podataka koristeći MS SQL Server kao primjer | 51 KB | |
| Istražite mogućnosti definiranja i korištenja prilagođenih tipova podataka, DML i DDL pokretača i ograničenja integriteta u postrelacijskom DBMS-u. Master metode i tehnologije za kreiranje prilagođenih tipova podataka na CLR.Net. | |||
| 73034. | Princip označavanja armiranobetonskih podnih ploča civilnih zgrada | 47,1 KB | |
| Princip označavanja armiranobetonskih podnih ploča civilnih zgrada U skladu sa GOST 2300978, podne ploče su označene brojnim alfanumeričkim indeksima. Prva grupa sadrži oznaku vrste ploče i njene projektne dimenzije, dužinu i širinu u decimetrima. | |||
Nervni sistem. Predavanje 2.
Bark Veliki mozak (cortex cerebri) je kontinuirani sloj sive tvari na površini moždanih hemisfera, debljine 2-5 mm. Nalazi se ne samo na zavojima, već se nastavlja u sve brazde, zbog čega je njegova površina relativno velika i iznosi 2200 cm 2 kod odrasle osobe. Masa kore je 581 g, zapremina oko 560 cm 3. U ovom slučaju, glavni volumen korteksa sastoji se od bijele tvari (450 cm 3, težina - 470 g). Telesna masa ćelija kortikalnih neurona je samo oko 21 g (20 cm3).
Veličina kortikalnih nervnih ćelija uveliko varira od 8-9 mk do 150 mk. U ljudskoj moždanoj kori postoji do 15 milijardi nervnih ćelija, od kojih su 6 milijardi male ćelije. Velika većina kortikalnih neurona pripada dvije vrste: piramidalnim neuronima i zvjezdanim neuronima. Ove ćelije se nalaze u korteksu u određenim slojevima.
U ljudskom mozgu postoji nekoliko filogenetski različitih različite vrste kora:
Drevni korteks - paleokorteks (0,6%) - praktički nema slojevitost (1-2 sloja), lokaliziran je u području olfaktornih trokuta, čini pokrivač septum pellucidum i okružuje jezgro amigdale.
Stari korteks - arhikorteks (2,2%) - ima 2-3 sloja, predstavljene sivom materijom dentatnog girusa, sivom materijom hipokampusa i medularnim strijama na corpus callosum.
Prijelazni korteks - mezokorteks (1,6%) - područje zasvođenog girusa.
Novi korteks - neokorteks (95,6%) - je dobro strukturiran i ima 6 slojeva.
Kod viših kralježnjaka pretežno je šest manje ili više jasno definiranih slojeva. Ali svaki od ovih slojeva, osim prvog, može se podijeliti na dva ili čak tri podsloja.
Prvi sloj, tzv. zonalni ili molekularna ploča, sastoji se uglavnom od pleksusa apikalnih dendrita piramidalnih neurona, čija se ćelijska tijela nalaze u drugim slojevima korteksa. U prvom sloju ima vrlo malo nervnih ćelija. Sadrži horizontalne ćelije čiji se aksoni i dendriti također nalaze horizontalno u istom sloju.
Drugi sloj, tzv. vanjski granularni ili vanjska granulirana ploča, uključuje masu malih ćelija koje pripadaju malim piramidalnim, takozvanim interneuronima i zvezdastim neuronima, a male piramidalne ćelije dominiraju u drugom sloju.
Treći sloj je vanjska piramidalna ploča sadrži piramidalne ćelije srednje veličine
Četvrti sloj, tzv. interni granularni ili unutrašnja granulirana ploča, sastoji se uglavnom od malih zvjezdanih ćelija, ali sadrži i male i srednje piramide.
Peti sloj je unutrašnja piramidalna ploča sadrži gigantske piramidalne ćelije ili Betzove ćelije.
Šesti sloj – multiformna ploča sadrže uglavnom piramidalne ćelije srednje veličine i mali broj malih piramidalnih i zvjezdastih ćelija.
Prva tri sloja su najmlađa i obezbeđuju vezu između različitih delova korteksa. Četvrti sloj je najrazvijeniji u područjima gdje pristižu aferentne informacije (osjetljivi centri, posebno postcentralni girus). Peti sloj je izražen u motoričkim područjima korteksa: precentralni girus, pericentralni lobuli, supramarginalni girus.
Svaki piramidalni neuron ima piramidalnu ćeliju i mnogo dendrita. Akson piramidalnog neurona izlazi iz malog brežuljka na dnu ćelije. U malim, takozvanim interkalarnim ili srednjim piramidalnim neuronima, aksoni, granajući se u horizontalnom ili vertikalnom smjeru, završavaju se odmah, bez napuštanja korteksa. Aksoni srednjih i velikih piramida daju mnoge kolaterale u korteksu, a glavna debla idu u subkortikalnu bijelu tvar. Neki od njih se vraćaju iz subkortikalne supstance u korteks date hemisfere ili, prolazeći kroz corpus callosum, završavaju u korteksu druge hemisfere. Služe za ujedinjavanje različitih dijelova moždane kore. Stoga se takvi piramidalni neuroni nazivaju asocijativni. Ostali aksoni su usmjereni na subkortikalne formacije i dalje na različite dijelove mozga i kičmene moždine. Ove piramide se zovu projekcija. U vezi sa filogenetskim razvojem sisara, broj piramidalnih ćelija se značajno povećava.
Širina čitavog korteksa, broj ćelija i širina svakog sloja ćelija u prednjem delu, kao i ćelijski sastav, odnosno veličina, oblik i raspored ćelija u svakom delu korteksa izuzetno variraju . Ove varijacije zauzimaju određene, jasno razgraničene teritorije. U mozgu velikih sisara postoji 11 jasno vidljivih velikih kortikalnih polja.
Na osnovu histoloških i fizioloških studija, svako takvo polje podijeljeno je u nekoliko nezavisnih sekcija. Značajnija diferencijacija korteksa uočena je kod viših sisara - majmuna. Posebno je sjajan kod ljudi.
Takvo istraživanje započeo je kijevski naučnik Vladimir Aleksejevič Bets, koji je 1874. godine objavio članak „Dva centra u moždanoj kori“, u kojem je opisao motoričku zonu u precentralnom girusu i osetljivu zonu u postcentralnom girusu.
Godine 1909. njemački neurolog Korbinian Brodmann objavio je karte citoarhitektonskih polja cerebralni korteks. Brodman je prvi napravio mape korteksa. Nakon toga, O. Vogt i C. Vogt (1919-1920), uzimajući u obzir strukturu vlakana, opisali su 150 mijeloarhitektonskih područja u moždanoj kori. Na Institutu za mozak Akademije medicinskih nauka SSSR-a, I. N. Filippov i S. A. Sarkisov kreirali su karte moždane kore, uključujući 47 citoarhitektonskih polja.
Podaci eksperimentalno istraživanje ukazuju da kada su određena područja moždane kore uništena ili uklonjena kod životinja, određene funkcije su poremećene. Ove činjenice su potvrđene klinička zapažanja za bolesne osobe s tumorskim lezijama ili ozljedama određenih područja moždane kore. Sve ovo nam je omogućilo da zaključimo da u kori velikog mozga postoje centri koji reguliraju obavljanje određenih funkcija. Morfološka potvrda fizioloških i kliničkih podataka bila je doktrina o različitoj kvaliteti strukture moždane kore u njenim različitim dijelovima – cito- i mijeloarhitektonici korteksa. Utvrđeno je da neuroni nisu locirani difuzno u korteksu, već su grupirani u ansamble.
Upotreba savremenih mikroelektrodnih metoda za proučavanje funkcija kortikalnih neurona značajno je proširila razumijevanje obrade senzornih informacija u neokorteksu i strukturne organizacije korteksa. Godine 1957. američki istraživač V. Mountcastle, analizirajući odgovore ćelija somatosenzornog (senzomotornog) korteksa mačke na podražaje različitih modaliteta, otkrio je sljedeće zanimljiva činjenica. Kada je mikroelektroda uronjena okomito na površinu somatosenzornog korteksa, sve ćelije na koje je naišla podjednako su reagovale na podražaj, na primer, na lagani dodir kože ili na pokret u zglobu. Ako je elektroda bila uronjena pod kutom u odnosu na površinu korteksa, tada su duž njenog puta dolazili neuroni s različitim senzornim modalitetima, naizmjenično s određenom periodičnošću.
Na osnovu ovih eksperimentalnih činjenica, W. Mountcastle je došao do zaključka da je somatosenzorni korteks organizovan u elementarne funkcionalne jedinice - stupovi orijentisan okomito na površinu. Prečnik takvog stuba je oko 500 μm određen horizontalnom distribucijom terminala aferentnog talamokortikalnog vlakna i vertikalnom orijentacijom dendrita piramidalne ćelije. Prema Mountcastleu, kolona je elementarni blok senzomotornog korteksa, gdje se vrši lokalna obrada informacija sa receptora jednog modaliteta. Ova hipoteza o stubnoj organizaciji neokorteksa postala je rasprostranjena i dala je podsticaj daljim istraživanjima u ovoj oblasti kod nas i u inostranstvu.
Prema moderne ideje, svaki funkcionalni stupac senzomotornog korteksa sastoji se od nekoliko morfoloških mikromodula koji ujedinjuju pet do šest neurona u obliku gnijezda. Ovaj modul uključuje nekoliko piramidalnih ćelija, čiji su apikalni dendriti što bliže i formiraju dendritski snop; unutar ovog snopa su mogući elektrotonske veze, koji osiguravaju, po svoj prilici, sinhroni rad cijele asocijacije.
Pored grupe vertikalno orijentisanih piramidalnih ćelija su zvezdaste ćelije, s kojim talamokortikalna vlakna dolaze u mikromodul u kontaktu. Neke od zvjezdastih stanica koje obavljaju inhibitornu funkciju imaju dugačke aksone koji se protežu horizontalno. Aksoni piramidalnih ćelija formiraju rekurentne kolaterale, koji mogu da obezbede i olakšavajuće uticaje unutar mikromodula i inhibitorne interakcije između mikromodula, kontaktirajući inhibitorne interneurone. Nekoliko strukturnih mikromodula, ujedinjenih horizontalnim grananjem završetaka specifičnih talamokortikalnih aferenata, aksona terminala i procesa zvjezdastih ćelija, formiraju stupac (ili makromodul), čiji promjer doseže 500-1000 µm. Svaki stupac karakterizira funkcionalno jedinstvo, koje se očituje u činjenici da neuroni kolone reagiraju na podražaj jednog modaliteta.
Nakon toga, princip stupne organizacije potvrđen je proučavanjem drugih projekcijske zone kora.
I.P. Pavlov je smatrao da je moždana kora neprekinuta perceptivna površina, kao skup kortikalnih krajeva analizatora. On je pokazao da kortikalni kraj analizatora nije neka strogo određena zona. Moždana kora je podijeljena na jezgro i raspršene elemente. Jezgro je mjesto koncentracije kortikalnih nervnih ćelija, koje čini tačnu projekciju svih elemenata određenog perifernog receptora, gdje se odvija veća analiza, sinteza i integracija funkcija. Raštrkani elementi mogu se nalaziti i duž periferije jezgra i na znatnoj udaljenosti od njega. Oni provode jednostavniju analizu i sintezu. Prisutnost raspršenih elemenata tokom destrukcije jezgre djelomično omogućava kompenzaciju narušene funkcije. Područja zauzeta raštrkanim elementima razni analizatori, mogu se naslagati jedna na drugu, preklapati jedna drugu. Dakle, moždana kora se može shematski zamisliti kao skup jezgara različitih analizatora, između kojih se nalaze razbacani elementi koji pripadaju različitim (susjednim) analizatorima. Gore navedeno nam omogućava da razgovaramo o tome dinamička lokalizacija funkcija u kori velikog mozga (I.P. Pavlov).
Razmotrimo položaj nekih kortikalnih krajeva različitih analizatora (nukleusa) u odnosu na vijuge i režnjeve moždanih hemisfera kod ljudi u skladu sa citoarhitektonskim mapama.
1. U korteksu postcentralnog girusa i gornjeg parijetalnog lobula leže nervne ćelije koje čine jezgro kortikalnog analizatora općeg osjetljivost(temperatura, bol, taktilna) i proprioceptivna. Senzorni putevi koji vode do kore velikog mozga se ukrštaju ili na nivou različitih segmenata kičmene moždine (putevi bola, temperaturne osetljivosti, dodira i pritiska) ili na nivou produžene moždine (putevi proprioceptivne osetljivosti kortikalnog pravca). Kao rezultat toga, postcentralni vijugi svake hemisfere su povezani sa suprotnom polovinom tijela. U postcentralnom girusu sva receptorska polja različitih dijelova ljudskog tijela su projektovana na način da se kortikalni krajevi analizatora osjetljivosti donjih dijelova tijela i donjih udova, a receptorska polja gornjih dijelova tijela i glave, i gornjih ekstremiteta su projektovana najniže (bliže lateralnom sulkusu).
2. Core motor analizator se nalazi uglavnom u takozvanom motornom korteksu, koji uključuje precentralni girus i paracentralni lobulu na medijalna površina hemisfere. U 5. sloju korteksa nalaze se precentralni girus piramidalni neuroni (Betz ćelije), koje je I.P. Pavlov klasifikovao kao interkalarne i primetio da su ove ćelije svojim procesima povezane sa subkortikalnim jezgrima, motornim ćelijama kranijalnih jezgara i kičmeni nervi. Štoviše, u gornjim dijelovima precentralnog girusa i u paracentralnom lobulu nalaze se ćelije, impulsi iz kojih se šalju u mišiće najnižih dijelova trupa i donjih ekstremiteta. U donjem dijelu precentralnog girusa nalaze se motorni centri koji reguliraju aktivnost mišića lica.
Tako su sva područja ljudskog tijela projektovana „naopačke” u precentralnom girusu. Zbog činjenice da se piramidalni putevi, koji potiču od gigantopiramidalnih ćelija, ukrštaju ili na nivou moždanog stabla (kortikonuklearna vlakna), na granici sa kičmenom moždinom, ili u segmentima kičmene moždine (kortikospinalni trakt), motor područja svake hemisfere su povezana sa skeletnim mišićima suprotne strane tijela. Ako su mišići udova izolovano povezani s jednom od hemisfera, mišići trupa, larinksa i ždrijela su povezani s motoričkim područjima obje hemisfere.
Motor I.P. Pavlov je područje moždane kore nazivao i područjem receptora, jer se ovdje radi o analizi proprioceptivnih (kinetičkih nadražaja koje opažaju receptori ugrađeni u skeletnih mišića, tetive, fascije i zglobne kapsule).
3. Jezgro analizatora pruža funkciju kombinovana rotacija glave i očiju unutra suprotnoj strani , je u stražnje regije srednji frontalni girus, u takozvanoj premotornoj zoni.
4. U predelu donjeg parijetalnog lobula, u supramarginalnom girusu, nalazi se jezgro motoričkog analizatora, čiji je funkcionalni značaj implementacija sinteza svima svrsishodna, profesionalna, radnička i sportska kretanja. Ovo jezgro je asimetrično. Za dešnjake je u lijevoj, a za ljevake- samo u desnoj hemisferi. Sposobnost koordinacije ovih složenih svrsishodnih pokreta pojedinac stječe tijekom života kao rezultat praktične aktivnosti i akumulacije iskustva. Namjerni pokreti nastaju zbog stvaranja privremenih veza između stanica smještenih u precentralnom i supramarginalnom vijuganju. Oštećenje polja ne uzrokuje paralizu, već samo dovodi do gubitka sposobnosti stvaranja složenih koordiniranih svrsishodnih pokreta - apraksija (praxis - praksa).
5. U korteksu gornjeg parijetalnog lobula nalazi se jezgro analizatora kože jedne od posebnih vrsta osjetljivosti, koja ima inherentnu funkciju prepoznavanje predmeta dodirom,- streognozija. Kortikalni kraj ovog analizatora nalazi se u desnoj hemisferi. Oštećenje površinskih slojeva korteksa u ovom dijelu je praćeno gubitkom funkcije prepoznavanja predmeta dodirom, iako su ostale vrste opće osjetljivosti očuvane.
6. U dubini lateralne brazde, na površini srednjeg dijela gornjeg temporalnog girusa okrenutog prema insuli (gdje je vidljiv transverzalni temporalni girus, ili Heschlov gyrus), nalazi se jezgro auditivni analizator. TO nervne celije, koji čini jezgro slušni analizator svaka od hemisfera, provodni putevi od receptora i lijeve i desna strana. S tim u vezi, jednostrano oštećenje ovog jezgra ne uzrokuje potpuni gubitak sposobnosti percepcije zvukova. Bilateralno oštećenje prati kortikalna gluvoća.
7. Core vizuelno analizator se nalazi u okcipitalni režanj moždane hemisfere u obodu kalkarinog sulkusa. Core vizuelni analizator desna hemisfera je povezana putevima sa lateralnom polovinom retine desnog oka i medijalnom polovinom retine lijevog oka. U korteksu okcipitalnog režnja lijeve hemisfere projiciraju se receptori lateralne polovine retine lijevog oka i medijalne polovice retine desnog oka. Kao i kod jezgra slušnog analizatora, samo bilateralno oštećenje jezgara vizuelnog analizatora dovodi do potpunog kortikalnog sljepila. Oštećenje polja koje se nalazi nešto iznad nukleusa vizuelnog analizatora je praćeno gubitkom vizuelne memorije, ali se gubitak vida ne primećuje. Još viši je dio korteksa, čiji je poraz popraćen gubitkom sposobnosti navigacije u nepoznatom okruženju.
8. Na donjoj površini temporalnog režnja hemisfere mozga, u predjelu uncusa i dijelom u području hipokampusa nalazi se jezgro olfaktorno analizator. Sa filogenetskog gledišta, ova područja pripadaju najstarijim dijelovima moždane kore. Osjeti mirisa i okusa su međusobno usko povezani, što se objašnjava bliskim položajem jezgara olfaktornog i okusnog analizatora. Također je primjećeno (V.M. Bekhterev) da je percepcija ukusa oštećena kada je oštećen korteks najnižih dijelova postcentralnog girusa. Jezgra analizatora ukusa i mirisa obe hemisfere povezana su sa receptorima na levoj i desnoj strani tela.
Opisani kortikalni krajevi nekih analizatora nisu prisutni u moždanoj kori samo ljudi, već i životinje. Specijalizovani su za percepciju, analizu i sintezu signala koji dolaze iz spoljašnjeg i unutrašnjeg okruženja, čineći, prema definiciji I. P. Pavlova, prvi signalni sistem stvarnosti.
Drugi sistem signalizacije postoji samo kod ljudi i određena je razvojem govora. Govor, a s njim i svijest, filogenetski su najmlađe funkcije mozga. U tom smislu, kortikalni krajevi analizatora su najmanje lokalizirani. Govorne i misaone funkcije izvode se uz sudjelovanje cijelog korteksa. Međutim, u kori velikog mozga moguće je razlikovati određena područja koja karakteriziraju strogo definirane govorne funkcije. Da, analizatori motoričkog govora(usmene i pismene) nalaze se pored motoričkog područja korteksa, tačnije, u onim dijelovima korteksa frontalnog režnja koji su uz precentralni girus.
Analizatori vizuelni i slušna percepcija govorni signali se nalaze pored analizatora vida i sluha. Treba istaći da su govorni analizatori kod dešnjaka lokalizovani u levoj hemisferi, a kod levorukih - uglavnom u desnoj.
Razmotrimo položaj nekih od govornih analizatora u moždanoj kori.
1. Motorni analizator pisanog govora nalazi se u zadnjem dijelu srednjeg frontalnog girusa. Djelatnost ovog analizatora usko je povezana s analizatorom svrsishodnih pokreta, koji osigurava formiranje naučenih pokreta ruku i očiju neophodnih za pisanje. Ako se ošteti, gubi se sposobnost izvođenja finih pokreta, koji se obično izvode pod kontrolom vida i neophodnih za crtanje slova, riječi i drugih znakova (agrafija).
2. Jezgro motornog analizatora artikulacija govora(govorno-motorni analizator) nalazi se u zadnjim dijelovima donjeg frontalnog girusa (Brokin centar). Oštećenje korteksa u ovom području dovodi do motoričke afazije, odnosno gubitka sposobnosti izgovaranja riječi (afazija). Ova afazija nije povezana s gubitkom sposobnosti kontrakcije mišića uključenih u proizvodnju govora. Štoviše, ako je ovo polje oštećeno, sposobnost izgovaranja zvukova ili pjevanja se ne gubi.
3. B centralna odjeljenja nalazi se donji frontalni girus jezgro govornog analizatora povezanog s pjevanjem. Njegov poraz prati vokalna amuzija - nesposobnost komponovanja i reprodukcije muzičkih fraza i agramatizam, kada se gubi sposobnost sastavljanja smislenih rečenica od pojedinačnih riječi. Govor takvih ljudi sastoji se od skupa riječi koje nisu povezane u semantičkom značenju.
4. Core slušni analizator govora blisko povezano sa
kortikalni centar slušnog analizatora i nalazi se u stražnjim dijelovima
gornji temporalni girus, na njegovoj površini okrenut u stranu
lateralni sulkus moždane hemisfere. Oštećenje nukleusa ne narušava slušnu percepciju zvukova općenito, ali se gubi sposobnost razumijevanja riječi i govora (verbalna gluvoća, ili senzorna afazija). Funkcija ovog jezgra je da osoba ne samo da čuje i razumije govor druge osobe, već i kontrolira svoj.
5. U srednjoj trećini gornjeg temporalnog girusa nalazi se jezgro kortikalnog analizatora, čiji je poraz praćen pojavom muzičke gluvoće, kada se muzičke fraze percipiraju kao besmislen skup raznih zvukova. Ovaj kortikalni kraj slušnog analizatora pripada centrima drugog signalnog sistema, koji percipiraju verbalno označavanje predmeta, radnji, pojava, tj. signali za detekciju signala.
6. U direktnoj vezi sa jezgrom vizuelnog analizatora nalazi se jezgro vizuelni analizator pisanog govora(polje 39), koji se nalazi u ugaonom girusu donjeg parijetalnog lobula. Oštećenje ovog nukleusa dovodi do gubitka sposobnosti percepcije pisanog teksta i čitanja (disleksija).
Ljudski frontalni korteks povezan je sa društvenim principom. Njegovi neuroni formiraju dvosmjerne veze sa svim ostalim područjima korteksa. Sa oštećenjem nastaju poremećaji društveno ponašanje, grubost, nelojalnost, smanjen osjećaj svrhe, apatija. Nakon lobotomije postoji potpuni nedostatak inicijative i društvene aktivnosti.
Asimetrija funkcija desne i lijeve hemisfere je vrlo izražena. Lijeva hemisfera daje logiku, apstraktno mišljenje, dominira u odnosu na govorne funkcije (čitanje, pisanje, brojanje) i složene voljne pokrete. Desna hemisfera– emocionalnost, imaginativno mišljenje, negovorne funkcije prepoznavanja složenih vizuelnih i slušnih slika, taktilna percepcija, percepcija prostora, oblika, pravca, intuitivno mišljenje.
Slični članci
-
Gdje možete vidjeti Mliječni put?
Fotografija sjevernog svjetla sa ISS-a Radi vaše udobnosti, napravili smo navigaciju kroz članak tako da možete brzo pronaći informacije koje su vam potrebne. Sjeverna (ili polarna) svjetlost, nebeski karneval, plesovi lisica, aurora borealis - ovo je prirodni fenomen...
-
Hidrogeologija, ili podzemne vode planete
Hidrogeologija je nauka o podzemnim vodama. Podzemne vode su one koje se nalaze ispod površine zemlje, ograničene na različite stijene i ispunjavaju pore, pukotine i kraške šupljine. Hidrogeologija proučava porijeklo i...
-
Jade Rod&Co (ljubavni biseri) Šta je štap od žada
Činjenica da izgledom podsjeća na dugačko deblo i zaobljenu glavu?.. Međutim, to ne sprječava ženu da sa radoznalošću ispituje muški penis, kao da ga upoznaje, a takođe je važna i kako izgleda. Nikada ne postoje dva, apsolutno...
-
Safiri iz Rusije: kako Monocrystal održava svjetsko vodstvo
JSC Monocrystal je najveći svjetski proizvođač sintetičkog safira i paste za metalizaciju solarnih ćelija. Safir, drugi najtvrđi kamen nakon dijamanta, ima odlične mehaničke, optičke i hemijske...
-
Kalendar dividendi Presjek za akcije Gazproma godišnje
Nastavkom korištenja naše stranice pristajete na obradu kolačića, korisničkih podataka (informacije o lokaciji, tip uređaja, IP adresa) u svrhu rada stranice, ponovnog ciljanja i...
-
Future Simple - jednostavno buduće vrijeme
Među oblicima vremena engleskog glagola, Future Simple vrijeme je jedno od najčešće korištenih. U kojim slučajevima se koristi jednostavno buduće vrijeme? Pogledajmo izbliza. Upotreba Future Simple Future vremena u...