Учени са преброили броя на невроните в човешкия мозък. Електрически прояви на активността на кората на главния мозък
Кората на полукълбата голям мозъке най-висшата и най-сложно организирана нервен центърекранен тип, чиято дейност осигурява регулирането на различни функции на тялото и сложни форми на поведение.
Кората е образувана от слой сиво вещество с дебелина 3-5 мм. Сивото вещество съдържа нервни клетки (повече от 10 милиарда неврони), нервни влакна и невроглиални клетки. Различните му части, които се различават една от друга по някои характеристики на местоположението и структурата на клетките, местоположението на влакната и функционалното значение, се наричат полета, които за първи път са описани от немския лекар и учен К. Бродман.
Цитоархитектоника
Сред многополярните неврони на кората се разграничават пирамидални, звездовидни, веретенообразни, паякообразни, хоризонтални, "канделаброви" клетки, клетки с двоен букет от дендрити и някои други видове неврони.
Пирамидални невронисъставляват основната и най-специфична форма за кората на главния мозък. Те имат удължено конусообразно тяло, чийто връх е обърнат към повърхността на кората. Дендритите се простират от върха и страничните повърхности на тялото. Аксоните произхождат от основата на пирамидалните клетки.
Пирамидалните клетки на различните слоеве на кората се различават по размер и имат различни функционална стойност. Малките клетки са интеркаларни неврони. Във формирането участват аксони на големи пирамиди моторпирамидални пътища.
Невроните на кората са разположени в неясно разграничени слоеве, които са обозначени с римски цифри и номерирани отвън навътре. Всеки слой се характеризира с преобладаването на който и да е тип клетки. Има шест основни слоя в мозъчната кора:
- I - молекулярен;
- II - външен гранулиран;
- III - пирамидален;
- IV - вътрешен гранулиран;
- V - ганглионен;
- VI - слой от полиморфни клетки.
аз- Молекулярнаслой кора съдържа голям броймалки асоциативни хоризонтални клетки на Cajal. Техните аксони вървят успоредно на повърхността на мозъка като част от тангенциалния плексус на нервните влакна на молекулярния слой. Въпреки това, по-голямата част от влакната на този плексус е представена от разклонения на дендритите на подлежащите слоеве.
II - външна гранулиранаслоят е образуван от множество малки пирамидални и звездовидни неврони. Дендритите на тези клетки се издигат в молекулярния слой, а аксоните или отиват в бели кахъри, или, образувайки дъги, също навлизат в тангенциалния плексус от влакна на молекулярния слой.
III - Най-широкият слой на мозъчната кора - пирамидален. Съдържа пирамидални неврони, клетки на Мартиноти и веретенообразни клетки с двоен букет от дендрити. Апикалните дендрити на пирамидите отиват в молекулярния слой, страничните дендрити образуват синапси със съседни клетки на този слой. Аксонът на пирамидната клетка винаги се отклонява от основата си. В малките клетки той остава в кората; в големите клетки образува миелиново влакно, което отива в бялото вещество на мозъка. Аксоните на малки полигонални клетки на Martinotti са насочени към молекулярния слой. Пирамидалният слой изпълнява предимно асоциативни функции.
IV- Вътрешно зърнистослой в някои области на кората е много силно развит (например в зрителната и слуховата кора), докато в други може да отсъства почти (например в прецентралната извивка). Този слой се образува от малки звездовидни неврони. Състои се от голям брой хоризонтални влакна.
V- Ганглийнислой на кората е образуван от големи пирамиди, а областта на моторната кора (прецентрален извивка) съдържа гигантски пирамиди , които за първи път са описани от киевския анатом V. A. Bets. Апикалните дендрити на пирамидите достигат до 1-ви слой. Аксоните на пирамидите се проектират към моторните ядра на мозъка и гръбначен мозък. Най-дългите аксони на клетките на Betz в пирамидните пътища достигат до каудалните сегменти на гръбначния мозък.
В допълнение към пирамидалните неврони в ганглиозния слой на кората има вертикални вретеновидни клетки, чиито аксони се издигат в I слойкора и кошничкови клетки.
VI - Слой полиморфни клеткиобразувани от неврони с различни форми (веретенообразни, звездовидни, клетки на Мартиноти). Аксоните на тези клетки преминават в бялото вещество като част от еферентните пътища, а дендритите достигат до молекулярния слой.
Миелоархитектоника
Сред нервните влакна на мозъчната кора могат да се разграничат асоциативенвлакна, които свързват отделни части на кората на едно полукълбо, комиссураленсвързване на кората на различни полукълба и проекциявлакна, аферентни и еферентни, които свързват кората с ядрата на долните части на централната нервна система. Проекционните влакна в кората на полукълбата образуват радиални лъчи, завършващи в III - пирамидален слой. В допълнение към вече описания тангенциален плексус на I - молекулярния слой, на нивото на IV - вътрешния гранулиран и V - ганглионен слой, има два тангенциални слоя от миелинизирани нервни влакна - съответно външната ивица на Баяргер и вътрешната ивица на Баяргер. Последните две системи са плексуси, образувани от крайните участъци на аферентните влакна.
Модулен принцип на кортикална организация
В кората на мозъчните полукълба са описани повтарящи се блокове или модули от неврони, които се считат за негови морфофункционални единици, способни на относително автономна дейност. Те имат формата на цилиндри или колони, преминаващи вертикално през цялата дебелина на кората.
Всеки модул включва аферентни пътища, система от локални връзки и еферентни пътища.
ДА СЕ аферентни пътищавключват кортико-кортикални и таламо-кортикални влакна.
Модулът е организиран около кортико-кортикални влакна, които са аксони на пирамидални клетки или в същото полукълбо, или в противоположното. Кортико-кортикалните влакна образуват окончания във всички слоеве на кората на даден модул.
Модулът също така включва таламо-кортикални влакна, които завършват в слой IV на кората на бодливи звездовидни неврони и базални дендрити на пирамидални неврони.
Еферентни пътищасе образуват от аксони на големи и гигантски пирамидални неврони, както и аксони на веретеновидни и някои други клетки от VI слой на кората.
Локална система за връзкисе формира от интеркаларните неврони на модула, които включват повече от дузина видове клетки. Повечето от тях са инхибиторни и регулират активността предимно на пирамидални неврони.
От инхибиторните неврони на модула най-висока стойностимам:
- аксо-аксонални клетки;
- клетки - "канделабри";
- кошничкови клетки;
- клетки с двоен букет от дендрити;
- клетки с аксонална четка.
Системата от инхибиторни неврони играе ролята на филтър, който инхибира част от пирамидалните неврони на кората.
Обвивки на главния и гръбначния мозък
Мозъкът и гръбначният мозък са покрити с три съединителнотъканни мембрани: мека, непосредствено съседна на мозъчните тъкани, арахноидна и твърда, която граничи с костна тъканчереп и гръбначен стълб. Менингите осъществяват защитна, вкл. амортисьорна функция, осигуряват производството и усвояването на цереброспиналната течност.
пиа матернепосредствено до мозъчната тъкан и ограничена от нея от маргиналната глиална мембрана. в рехави влакнести съединителната тъканмембрани има голям брой кръвоносни съдове, които захранват мозъка, множество нервни влакна, терминален апарат и единични нервни клетки. Pia mater обгражда съдовете, проникващи в мозъка, образувайки периваскуларна пиална мембрана около тях. Във вентрикулите на мозъка пиа матер, заедно с епендимата, участват в образуването на хороидни плексуси, които произвеждат цереброспинална течност.
Арахноидаленпредставена от тънък слой рехава фиброзна съединителна тъкан. Между него и пиа матер лежи мрежа от напречни греди, състояща се от тънки снопчета колаген и тънки еластични влакна. Тази мрежа свързва черупките заедно. Между пиа матер, която повтаря релефа на мозъчната тъкан, и арахноида, преминаващ през издигнатите области, без да навлиза в вдлъбнатините, има субарахноидно (субарахноидно) пространство, пронизано с тънък колаген и еластични влакна, които свързват мембраните помежду си. Субарахноидалното пространство комуникира с вентрикулите на мозъка и съдържа цереброспинална течност. Тази област е домакин на големи кръвоносни съдове, чиито клони хранят мозъка.
Власинките на арахноидната мембрана (най-големите се наричат пахионични гранулации) служат като места, през които веществата от цереброспиналната течност се връщат в кръвта. Те са аваскуларни израстъци на арахноида, съдържащи мрежа от подобни на прорези пространства и изпъкнали в лумена на синусите на твърдата мозъчна обвивка.
Дура матеробразувана от плътна фиброзна съединителна тъкан, съдържаща много еластични влакна. В черепната кухина тя е плътно слята с периоста. IN гръбначния каналтвърдата мозъчна обвивка е ограничена от гръбначния периост от епидурално пространство, изпълнено със слой от рехава фиброзна съединителна тъкан, което му осигурява известна подвижност. Между твърдата мозъчна обвивка и арахноида е субдуралното пространство. Субдуралното пространство съдържа малко количество течност.
Мембраните от страната на субдуралното и субарахноидалното пространство са покрити със слой плоски клеткиглиална природа.
Възрастови промени
Промени в централната нервна система при старостсвързани предимно със склеротични промени в мозъчните съдове. В напреднала възраст меките и арахноидни мембрани на мозъка се удебеляват. В тях може да се появят варовикови отлагания. Има атрофия на кората полукълбаглавно във фронталните и париеталните дялове. Броят на невроните на единица обем на кората намалява, това зависи главно от клетъчната смърт. Невроните намаляват по размер, частично губят базофилната си субстанция, ядрата стават по-плътни, очертанията им стават неравномерни. Пирамидите на V слой на моторната кора и крушовидните клетки на кората на малкия мозък се променят по-бързо от останалите. Липофусциновите гранули се натрупват в неврони на различни части на нервната система.
Всички най-важни поведенчески реакции на човека се осъществяват с помощта на централната нервна система. Основните функции на ЦНС са:
- интеграция на всички части на тялото в едно цяло и тяхното регулиране;
- управление на състоянието и поведението организъм в съответствие с условията външна средаи неговите нужди.
При висшите животни и човека водещата част на ЦНС е кората на главния мозък . Той контролира най-сложните функции в човешкия живот умствени процеси(съзнание, мислене, реч, памет и др.).
Основните методи за изследване на функциите на централната нервна система са методите за отстраняване и дразнене (в клиниката и върху животни), регистрация на електрически явления, методът условни рефлекси.
Продължават да се разработват нови методи за изследване на централната нервна система: с помощта на т.нар компютърна томографияможете да видите морфофункционалните промени в мозъка на различни дълбочини; фотография в инфрачервени лъчи (термично изображение) ви позволява да откриете най-горещите точки на мозъка; нови данни за работата на мозъка дава изследването на неговите магнитни трептения.
Основни функции и взаимодействия на невроните
Основните структурни елементи на нервната система са нервните клетки или невроните.
Основни функции на невроните
Чрез невроните информацията се предава от една част на нервната система към друга, обменя се информация между нервната система и различни части на тялото. Невроните извършват сложни процеси на обработка на информация. С тяхна помощ, отговориорганизма (рефлекси) към външни и вътрешни стимули.
По този начин, основните функции на невроните са: възприемане на външни стимули рецепторна функция, обработката им интегративна функция и предаване на нервни влияния към други неврони или различни работни органи ефекторна функция.В тялото на нервната клетка, или сома, протичат основните процеси на обработка на информацията. Многобройни дървовидни разклонени процеси дендрити(гръцки dendron дърво) служат като невронни входове, през които сигналите влизат в нервната клетка. Изходът на неврона е процес, който се простира от тялото на клетката аксон(гръцки axis ос), която предава нервните импулси по-нататък към друга нервна клетка или работен орган (мускул, жлеза). Особено висока възбудимост има началната част на аксона и разширението на мястото на излизането му от тялото на клетката. аксонов хълмневрон. Именно в този сегмент на клетката възниква нервният импулс.
Видове неврони
Невроните се делят на три основни типа: аферентни, еферентни и междинни. Аферентни неврони(чувствителни или центростремителни) предават информация от рецепторите към 1CNS. Телата на тези неврони са разположени извън ЦНС в гръбначни възлии във възли черепномозъчни нерви. Аферентните неврони имат дълъг процес - дендрит, който контактува по периферията с възприемащо образувание - рецептор или сам образува рецептор, както и втори процес - аксон, който влиза през задни рогав гръбначния мозък.
Еферентни неврони(центробежни) са свързани с прехвърлянето на низходящи влияния от горните етажи на нервната система към подлежащите или от централната нервна система към работните органи. Еферентните неврони се характеризират с разклонена мрежа от къси процеси - дендрити и един дълъг процес - аксон.
междинни неврони(интернейрони или интеркаларни) - това са, като правило, по-малки клетки, които комуникират между различни (по-специално аферентни и еферентни) неврони. Те предават нервни въздействия в хоризонтална посока (например в рамките на един сегмент на гръбначния мозък) и във вертикална посока (например от един сегмент на гръбначния мозък към други - по-високи или долни сегменти). Благодарение на многобройните разклонения на аксона, междинните неврони могат едновременно да възбуждат голям брой други неврони.
Възбудни и инхибиторни синапси
Взаимодействието на невроните един с друг (и с ефекторни органи) се осъществява чрез специално образованиесинапси (гръцки контакт). Те се образуват от крайни клонове на неврон върху тялото или процеси на друг неврон. Колкото повече синапси има върху една нервна клетка, толкова повече тя възприема различни стимули и следователно толкова по-широка е сферата на влияние върху нейната дейност и възможността за участие в различни реакции на тялото. Особено много синапси има във висшите части на нервната система и то в невроните с най-сложни функции.
В структурата на синапса се разграничават три елемента (фиг. 1):
- пресинаптична мембрана, образуван от удебеляване на мембраната на крайния клон на аксона;
- синаптична цепнатинамежду неврони;
- постсинаптична мембранаудебеляване на съседната повърхност на следващия неврон.
Ориз. 1. Диаграма на синапс
Пред. пресинаптична мембрана,
Бърз. постсинаптична мембрана,
C синоптични мехурчета,
Щ синоптична празнина,
М митохондрии,
Ах ацетилхолин
Р рецептори и пори (Пори)
дендрит (D) следващ
неврон.
Стрелка едностранно провеждане на възбуждане.
В повечето случаи прехвърлянето на влиянието на един неврон върху друг се извършва химически. Пресинаптичната част на контакта съдържа синаптични везикули които съдържат специални вещества посреднициили посредници. Те могат да бъдат ацетилхолин (в някои клетки на гръбначния мозък, във автономните възли), норепинефрин (в окончанията на симпатиковите нервни влакна, в хипоталамуса), някои аминокиселини и др.. Нервните импулси, пристигащи в края на аксона, причиняват изпразването на синаптичните везикули и екскрецията на медиатора в синаптичната цепнатина.
По естеството на въздействието върху последващата нервна клетка се разграничават възбуждащи и инхибиторни синапси.
При възбудни синапсимедиатори (например ацетилхолин) се свързват със специфични макромолекули на постсинаптичната мембрана и причиняват нейната деполяризация. В този случай малка и краткотрайна (около 1 ms) флуктуация на мембранния потенциал към деполяризация или възбуден постсинаптичен потенциал(VPSP). За да възбуди неврон, е необходимо EPSP да достигне прагово ниво. За това големината на деполяризационното изместване на мембранния потенциал трябва да бъде най-малко 10 mV. Действието на медиатора е много кратко (1-2 ms), след което се разделя на неефективни компоненти (напр. ацетилхолинът се разгражда от ензима холинестераза до холин и оцетна киселина ) тиня и се реабсорбира обратно от пресинаптичните окончания (например норепинефрин).
IN инхибиторни синапсисъдържащи се спирачки (Например, гама-аминомаслена киселина ). Тяхното действие върху постсинаптичната мембрана предизвиква увеличаване на освобождаването на калиеви йони от клетката и увеличаване на поляризацията на мембраната. В този случай се записва краткотрайна флуктуация на мембранния потенциал в посока хиперполяризация инхибиторен постсинаптичен потенциал(TPSP). В резултат на това нервната клетка се инхибира. По-трудно е да го възбудите, отколкото в първоначалното състояние. Това ще изисква по-силна стимулация за достигане на критично ниво на деполяризация.
Появата на импулсен отговор на неврон
Както възбудните, така и инхибиторните синапси са разположени върху мембраната на тялото и дендритите на нервната клетка. . В определени моменти от време някои от тях могат да бъдат неактивни, докато другата част има активно въздействие върху мембранните зони, съседни на тях. Цялостната промяна в мембранния потенциал на неврона е резултат от сложно взаимодействие(интегриране) на локални EPSP и IPSP на всички многобройни активирани синапси. При едновременното въздействие както на възбуждащите, така и на инхибиторните синапси се получава алгебрично сумиране (т.е. взаимно изваждане) на техните ефекти. В този случай възбуждането на неврона ще настъпи само ако сумата от възбудни постсинаптични потенциали ще бъде повече от суматаинхибиторен . Това превишение трябва да бъде определена прагова стойност (около 10 mV). Само в този случай се проявява потенциалът за действие на клетката. Трябва да се отбележи, че като цяло възбудимостта на неврона зависи от неговия размер: колкото по-малка е клетката, толкова по-висока е нейната възбудимост .
С появата на потенциал за действие, процесът на провеждане нервен импулспо аксона и го прехвърля към следващия неврон или работен орган, т.е. осъществява се ефекторната функция на неврона. Нервният импулс е основното средство за комуникация между невроните.
По този начин, предаването на информация в нервната система става чрез два електрически механизма (EPSP; IPSP; потенциал за действие) и химически (медиатори).
Характеристики на дейността на нервните центрове
Свойствата на нервните центрове са до голяма степен свързани с характеристиките на провеждането на нервните импулси през синапсите, които свързват различни нервни клетки.
Характеристики на провеждането на възбуждане през нервните центрове
нервен център нарича съвкупността от нервни клетки, необходими за изпълнението на всяка функция. Тези центрове отговарят с подходящи рефлексни реакции на външна стимулация от свързаните с тях рецептори. Клетките на нервните центрове реагират и на прякото им дразнене от вещества в кръвта, протичаща през тях (хуморални влияния). В целия организъм има строга координация - координация на тяхната дейност.
Провеждането на вълна на възбуждане от един неврон към друг през синапса се осъществява в повечето нервни клетки по химически начинс помощта на медиатор, а медиаторът се съдържа само в пресинаптичната част на синапса и отсъства в постсинаптичната мембрана. Ето защо важна характеристикапровеждане на възбуждане чрез синоптични контакти еедностранно задържаненервни влияния, което е възможно само от пресинаптичната мембрана към постсинаптичната и е невъзможно в обратна посока. В тази връзка потокът от нервни импулси в рефлексната дъга има определена посока от аферентни неврони към интеркаларни и след това към еферентни мотоневрони или автономни неврони.
Голямо значениев дейността на нервната система има друга особеност на провеждане на възбуждане през синапсибавна проводимост.Времето, прекарано в процесите, протичащи от момента, в който нервният импулс се приближи до пресинаптичната мембрана, докато потенциалите се появят в постсинаптичната мембрана, се нарича синаптично забавяне. В повечето централни неврони тя е около 0,3 ms. След това е необходимо повече време за развитието на възбудния постсинаптичен потенциал (EPSP) и потенциала за действие. Целият процес на предаване на нервен импулс (от акционния потенциал на една клетка до акционния потенциал на следващата клетка) през един синапс отнема приблизително 1,5 ms. При умора, охлаждане и редица други влияния продължителността на синаптичното забавяне се увеличава. Ако за осъществяването на някаква реакция е необходимо участие Голям бройневрони (много стотици и дори хиляди), тогава общото забавяне на проводимостта през нервните центрове може да бъде десети от секундата и дори цели секунди.
С рефлекторно действие общо времеот момента на прилагане на външен дразнител до появата на отговор на организма – т.нарлатентно или латентно рефлексно времесе определя основно от продължителността на провеждане през синапсите. Стойността на латентното време на рефлекса играе важна роля индикатор функционално състояниенервни центрове . Измерването на латентното време на проста човешка двигателна реакция към външен сигнал се използва широко в практиката за оценка на функционалното състояние на ЦНС (фиг. 2).
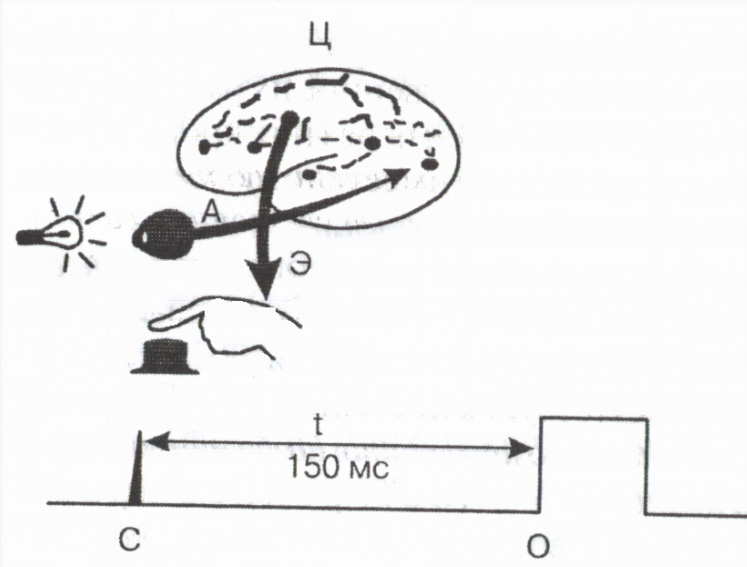
Ориз. 2. Схема за измерване на времето на двигателна реакция
Аферент
E еферентни и C централни пътища; C - светлинна сигнална маркировка,
О знак за натискане на бутон,
t 150ms време за реакция.
Сумиране на възбуждане
В отговор на единична аферентна вълна от рецептори към неврони, малко количество медиатор се освобождава в пресинаптичната част на синапса. В този случай EPSP обикновено се появява в постсинаптичната мембрана на неврона - малка локална деполяризация. За да може общата стойност на EPSP за цялата невронна мембрана да достигне прага за възникване на потенциал за действие, е необходимо сумиране на много подпрагови EPSP върху клетъчната мембрана. Само в резултат на такова сумиране на възбуждане невронът реагира. Правете разлика между пространствено и времево сумиране.
Пространствено сумираненаблюдава се в случай на едновременно получаване на няколко импулса в един и същ неврон през различни пресинаптични влакна. Едновременното възбуждане на синапсите в различни части на невронната мембрана увеличава амплитудата на общия EPSP до прагова стойност. В резултат на това възниква импулс за отговор на неврона и се осъществява рефлексна реакция. Например, получаването на отговор на двигателните клетки на гръбначния мозък обикновено изисква едновременно активиране на 50-100 аферентни влакна от съответните периферни рецептори.
Времево сумираневъзниква, когато един и същ аферентен път се активира от серия от последователни стимули. Ако интервалите между входящите импулси са достатъчно кратки, EPSP на неврона от предишни стимули нямат време да избледнеят, тогава следващите EPSP се наслагват един върху друг, докато деполяризацията на невронната мембрана достигне критично ниво за възникване на потенциал за действие. По този начин дори слаби раздразненияслед известно време те могат да предизвикат реакции на тялото (например кихане и кашляне в отговор на леки дразнения на лигавицата на дихателните пътища).
Трансформация и асимилация на ритъма
Естеството на отговора на неврона зависи не само от свойствата на стимула, но и от функционалното състояние на самия неврон.(неговия мембранен заряд, възбудимост, лабилност). Нервните клетки имат способността да променят честотата на предаваните импулси, т.е. свойство на трансформация на ритъм.
При висока възбудимост на неврон (например след прием на кофеин), повишена импулсация (умножаване на ритъм) и с ниска възбудимост (например с умора) забавяне на ритъма , тъй като трябва да се сумират няколко входящи импулса, за да се достигне накрая прага за поява на потенциал за действие. Тези промени в честотата на импулсите могат да засилят или отслабят реакциите на тялото към външни стимули.
С ритмични стимули активността на неврона може да се настрои към ритъма на входящите импулси, т.е. феномен на научаване на ритъм(Ухтомски А.А., 1928 г.). Развитието на асимилацията на ритъма осигурява настройка на дейността на много нервни центрове при управлението на сложни двигателни действия е особено важно да се поддържа темпото на цикличните упражнения.
следи процеси
След края на действието на стимула активното състояние на нервната клетка или нервния център обикновено продължава известно време. Продължителността на следовите процеси е различна: кратка в гръбначния мозък (няколко секунди или минути), много по-голяма в центровете на мозъка (десетки минути, часове или дори дни) и много дълга в мозъчната кора (до няколко десетилетия).
Импулсите, циркулиращи през затворени вериги от неврони, могат да поддържат ясно и краткотрайно състояние на възбуда в нервния център. Дългосрочните скрити следи са много по-трудни по природа. Предполага се, че дълготрайното запазване на следи в нервната клетка с всички характерни свойствастимулът се основава на промяна в структурата на протеините, които изграждат клетката, и на преструктурирането на синаптичните контакти.
Кратките импулсни последействия (с продължителност до 1 час) са в основата на т.нар краткотрайна памет , и дългосрочни следи, свързани със структурни и биохимични пренареждания в клетките, са в основата на образуването дългосрочна памет.
Координация на ЦНС
Процесите на координация на дейността на централната нервна система се основават на координацията на два нервни процеса - възбуждане и инхибиране. Инхибирането е активен нервен процес , което предотвратява или потиска възбудата.
Стойността на процеса на инхибиране в централната нервна система
Феноменът на инхибиране в нервните центрове е открит за първи път от И. М. Сеченов през 1862 г. Значението на този процес е разгледано от него в книгата "Рефлекси на мозъка" (1863 г.).
Като потопите жабешкия бут в киселина и същевременно раздразните някои части на мозъка (например като нанесете кристал готварска солв областта на диенцефалона), И. М. Сеченов наблюдава рязко забавяне и дори пълно отсъствие на "киселинния" рефлекс на гръбначния мозък (отдръпване на лапата). Следователно той заключава, че някои нервни центрове могат значително да променят рефлексната активност в други центрове, по-специално, горните нервни центрове могат да инхибират дейността на основните. Описаното преживяване влезе в историята на физиологията под името Инхибиране на Сеченов .
Инхибиторните процеси са необходим компонент на координацията на нервната дейност. първо, процесът на инхибиране ограничава разпространението на възбуждането до съседни нервни центрове, което допринася за концентрацията му в необходимите части на нервната система. Второ, възниквайки в някои нервни центрове успоредно с възбуждането на други нервни центрове, процесът на инхибиране по този начин изключва ненужните дейности този моменттела . Трето, развитието на инхибиране в нервните центрове ги предпазва от прекомерно пренапрежение по време на работа, тоест играе защитна роля.
Постсинаптично и пресинаптично инхибиране
Процес на спиране , за разлика от възбуждането, не може да се разпространява по нервното влакно винаги е локален процес в областта на синаптичните контакти. Според мястото на възникване се разграничават пресинаптично и постсинаптично инхибиране.
Постсинаптично инхибиранетова са инхибиторни ефекти, възникващи в постсинаптичната мембрана. Най-често този тип инхибиране се свързва с наличието в централната нервна система на специални инхибиторни неврони. Те са специален тип интеркаларни неврони, в които аксонните окончания освобождават инхибиторен невротрансмитер. Един от тези посредници е гама-аминомаслена киселина (GAM K).
Нервните импулси, приближаващи инхибиторните неврони, предизвикват в тях същия процес на възбуждане, както в другите нервни клетки.В отговор обичайният потенциал на действие се разпространява по аксона на инхибиторната клетка. Въпреки това, за разлика от други неврони, окончанията на аксоните освобождават не възбуждащ, а инхибиторен невротрансмитер. В резултат на това инхибиторните клетки инхибират онези неврони, на които завършват техните аксони.
Специалните инхибиторни неврони включват клетки на Раншоу в гръбначния мозък, клетки на Пуркиние в малкия мозък, кошничкови клетки в диенцефалона и др.Например, инхибиторните клетки са от голямо значение за регулиране на активността на мускулите-антагонисти: като водят до отпускане на мускулите-антагонисти, те по този начин улесняват едновременното свиване на мускулите-антагонисти (фиг. 3).
Клетки на Реншоуучастват в регулирането на нивото на активност на отделните моторни неврони на гръбначния мозък. Когато моторният неврон е възбуден, импулсите се движат по неговия аксон към мускулните влакна и едновременно по колатералите на аксона към инхибиторната клетка на Раншоу. Аксоните на последния се "връщат" към същия неврон, причинявайки неговото инхибиране. Колкото повече възбудни импулси изпраща моторният неврон към периферията (и оттам към инхибиторната клетка), толкова по-силен е този обратно спиране(вид постсинаптично инхибиране). Такива затворена системадейства като механизъм за саморегулиране на неврона, предпазвайки го от прекомерна активност.
Клетки на Пуркиние на малкия мозъктехните инхибиторни ефекти върху клетките на подкоровите ядра и стволовите структури участват в регулацията на мускулния тонус.
кошничкови клеткив диенцефалона те са, така да се каже, врати, които позволяват или не пропускат импулси, отиващи към мозъчната кора от различни области на тялото.
пресинаптично инхибираневъзниква преди синаптичния контакт в пресинатичния регион. Краят на аксона на инхибиторната нервна клетка образува синапс в края на аксона на възбудителната нервна клетка, причинявайки прекалено силна деполяризация на мембраната на този аксон, което инхибира преминаващите тук потенциали на действие и по този начин блокира предаването на възбуждане. Този тип инхибиране ограничава потока от аферентни импулси към нервните центрове, изключвайки влияния, които са външни за основната дейност.
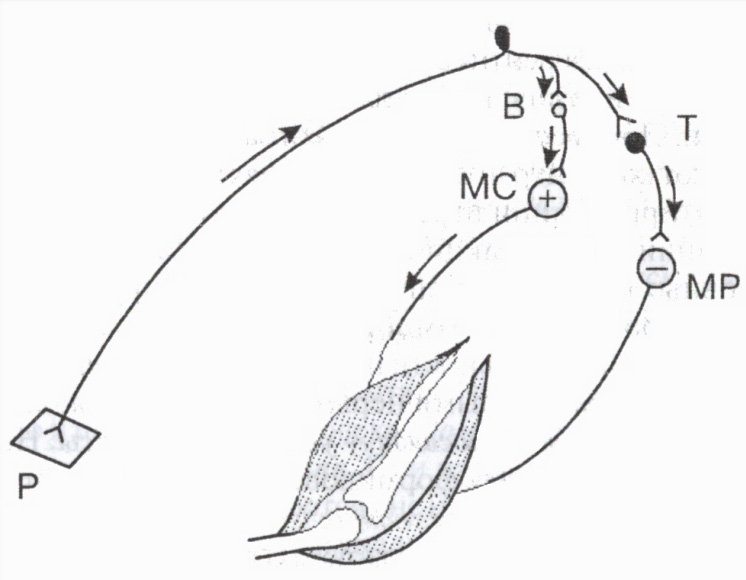
Ориз. 3. Участие на инхибиторната клетка в регулацията на мускулите антагонисти
В и Т - възбудни и инхибиторни неврони. Възбуждане (+) моторен неврон на мускула флексор (MS) и инхибиране (-) двигателен неврон на мускул екстензор (MR). P кожен рецептор.
Феноменът на облъчване и концентрация
Когато един рецептор се стимулира, възбуждането може по принцип да се разпространи в ЦНС във всяка посока и към всяка нервна клетка. Това се дължи на многобройните взаимовръзки на невроните на един рефлексна дъгас неврони на други рефлексни дъги. Разпространението на процеса на възбуждане към други нервни центрове се нарича феноменоблъчване.
Колкото по-силна е аферентната стимулация и колкото по-висока е възбудимостта на околните неврони, толкова повече неврони са обхванати от процеса на облъчване. Процесите на инхибиране ограничават облъчването и допринасят за концентрацията на възбуждане в началната точка на ЦНС.
Процесът на облъчване играе важна роля положителна роляпри формирането на нови реакции на тялото (ориентиращи реакции, условни рефлекси). Колкото повече различни нервни центрове се активират, толкова по-лесно е да се изберат сред тях центровете, които са най-необходими за последваща дейност. Благодарение на облъчването възбуждане между различни нервни центрове нови функционални отношения възникват условни рефлекси . На тази основа е възможно например да се формират нови двигателни умения.
В същото време облъчването на възбуждането също може отрицателно въздействие върху състоянието и поведението на тялото, нарушавайки фините връзки между възбудени и инхибирани нервни центрове и причинявайки нарушения на координацията на движението .
Доминантен
Изследвайки характеристиките на междуцентралните отношения, А. А. Ухтомски откри, че ако в животинското тяло възникне сложна рефлекторна реакция, например повтарящи се актове на преглъщане, тогава електрическата стимулация на двигателните центрове не само престава да предизвиква движение на крайниците в този момент, но и засилва протичането на верижната реакция, която е започнала, преглъщането, което се оказа доминиращо.
Такъв доминиращ фокус на възбуждане в централната нервна система, който определя текущата активност на организма, е обозначен от AA Ukhtomsky (1.923) с термина доминиращ.
Доминиращ фокус може да възникне, когато повишено нивовъзбудимост на нервните клетки, която се създава от различни хуморални и нервни влияния. Той потиска активността на други центрове, осигурявайки конюгирано инхибиране.
Обединяването на голям брой неврони в една доминираща система става чрез взаимно приспособяване към общия темп на дейност, т.е. чрез усвояване на ритъма. Някои нервни клетки забавят по-високата си скорост на активност, докато други увеличават ниската си скорост до някакъв среден, оптимален ритъм. Доминантата може да остане дълго време в скрито, следово състояние (потенциална доминанта). Когато се възобнови предишното състояние или предишната външна ситуация, доминантата може да се появи отново (актуализация на доминантата). Например, в предстартовото състояние се активират всички онези нервни центрове, които са били част от работната система по време на предишни обучения, и съответно се подобряват функциите, свързани с работата. умствено изпълнение упражнениеили представянето на движенията също възпроизвежда работната доминанта, която осигурява тренировъчния ефект на представянето на движенията и е в основата на така нареченото идеомоторно обучение. Когато сте напълно отпуснати (напр автогенен тренинг) спортистите постигат елиминиране на работещите доминанти, което ускорява процеса на възстановяване.
Като фактор на поведението доминантата се свързва с висшето нервна дейности човешката психология. Доминантно е физиологична основаакт на внимание. При наличието на доминанта много влияния на външната среда остават извън вниманието ни, но тези, които са особено интересни за нас, се улавят и анализират по-интензивно. По този начин доминантата е мощен фактор при избора на биологично и социално най-значимите стимули.
Функции на гръбначния мозък и подкоровите области на мозъка
В ЦНС, по-древни сегментен и еволюционно по-млад надсегментен части на нервната система. Сегментните отдели включват гръбначния, продълговатия и средния мозък, чиито отдели регулират функциите на отделни части на тялото, разположени на едно ниво. Супрасегментните отдели на диенцефалона, малкия мозък и кората на главния мозък нямат пряка връзка с органите на тялото, но контролират дейността си чрез подлежащите сегментни дялове.
Гръбначен мозък
Гръбначният мозък е най-долната и най-древна част от ЦНС. . Сивото вещество на човешкия гръбначен мозък съдържа около 13,5 милиона нервни клетки. От тях по-голямата част (97%) са междинни клетки (интернейрони или интернейрони) , които осигуряват сложни координационни процеси в гръбначния мозък. Сред двигателните неврони на гръбначния мозък, голям алфа моторни невронии малки гама моторни неврони. Най-дебелите и най-бързо проводящи влакна на моторните нерви се отклоняват от алфа моторните неврони, причинявайки съкращенияскелетна мускулни влакна. Тънките влакна на гама моторните неврони не предизвикват мускулна контракция. Те пасват на проприорецепторите мускулни вретенаи регулирайте тяхната чувствителност.
Рефлекси на гръбначния мозъкмогат да се подразделят на мотор извършва се от алфа моторните неврони на предните рога и вегетативен извършва се от аферентни клетки на страничните рога.
Моторните неврони на гръбначния мозък инервират всички скелетни мускули (с изключение на мускулите на лицето) . Гръбначният мозък осъществява елементарни моторни рефлексифлексия и екстензор, ритмичен, стъпващ, възникващ от дразнене на кожата или проприорецепторите на мускулите и сухожилията, а също така изпраща постоянни импулси към мускулите, поддържащи мускулен тонус. Специални двигателни неврони инервират дихателната мускулатура - междуребрените мускули и диафрагмата и осигуряват дихателни движения. Автономните неврони инервират всички вътрешни органи (сърце, кръвоносни съдове, потни жлези, жлези вътрешна секреция, храносмилателен тракт, пикочна система).
Функция на проводникана гръбначния мозък е свързано с предаването на по-горните части на нервната система на потока от информация, получен от периферията, и с провеждането на импулси, идващи от мозъка към гръбначния мозък.
Отзад последните годиниразвити специални техникиза изследване на дейността на гръбначния мозък в здрав човек. Например, функционалното състояние на алфа моторните неврони се оценява чрез промени в отговорните потенциали на мускулите по време на периферни стимули, т.нар. H-рефлекс (рефлекс на Хофман) мускул на прасецапри раздразнение тибиален нерви от Т-рефлекс (от tendon tendon) солеус мускул с дразнене на ахилесовото сухожилие. Разработени са техники за записване (от непокътнати кожи на тялото) на потенциали, преминаващи през гръбначния мозък към мозъка.
продълговатия мозък и варолиевия мост
Продълговатият мозък и мостът (най-общо задният мозък) са част от мозъчния ствол. Тук се намира голяма група черепномозъчни нерви (от V до XII чифт), които инервират кожата, лигавиците, мускулите на главата и редица вътрешни органи(сърце, бял дроб, черен дроб). Тук се намират центрове на много храносмилателни рефлекси дъвчене, преглъщане, движения на стомаха и част от червата, секреция на храносмилателни сокове и центрове на някои защитни рефлекси (кихане, кашляне, мигане, сълзене, повръщане) и центрове на водно-солевия и захарен метаболизъм . В дъното на IV вентрикула в продълговатия мозък е жизненоважен дихателен център, състоящ се от центрове на вдишване и издишване. Състои се от малки клетки, които изпращат импулси към дихателните мускули чрез двигателните неврони на гръбначния мозък.
Намира се в непосредствена близост сърдечно-съдов център . Неговите големи клетки регулират дейността на сърцето и лумена на кръвоносните съдове. Преплитането на клетките на дихателния и сърдечно-съдовия център осигурява тясното им взаимодействие.
Продълговатият мозък играе важна роля в осъществяването на двигателните актове и в регулирането на тонуса на скелетните мускули, повишаване на тонуса на екстензорните мускули . Той участва по-специално в изпълнението постурални рефлекси (цервикален, лабиринтен). Възходящо преминава през продълговатия мозък слухови, вестибуларни, проприоцептивни и тактилни пътища .
среден мозък
Средният мозък съдържа квадригемината, субстанцията нигра и червените ядра. В предните туберкули на квадригемината са зрителни подкорови центрове , и в задната част слухови . Междинният мозък участва в регулиране на движенията на очите , извършва зеничен рефлекс (разширение на зениците на тъмно и свиване на светло).
Квадригемината извършва поредица от реакции, които са Компоненти на ориентировъчния рефлекс . В отговор на внезапно дразнене главата и очите се обръщат към стимула, а при животните ушите са нащрек. Този рефлекс (според I. P. Pavlov, рефлексът „Какво е това?“) е необходим, за да подготви тялото за своевременна реакция към всяко ново въздействие.
черно вещество средният мозък е свързан с рефлексите на дъвчене и преглъщане, участва в регулиране на мускулния тонус (особено когато правите малки движенияпръстите) и в организирането на приятелски двигателни реакции.
червено ядро средният мозък изпълнява двигателни функции регулира тонуса на скелетната мускулатура , което води до повишаване на тонуса на мускулите флексори. Имайки значителен ефект върху тонуса на скелетните мускули, средният мозък участва в редица регулиране на рефлексите за поддържане на позата (коригираща настройка на тялото с темето на главата нагоре и т.н.).
диенцефалон
Диенцефалонът се състои от таламуса (зрителни туберкули) и хипоталамуса (хипоталамуса).
Презталамус преминават през всички аферентни пътища (с изключение на обонятелните), които се изпращат до съответните възприемащи области на кората (слухови, зрителни и др.). Ядрата на таламуса се делят на специфични и неспецифични . Конкретните са комутационни (релейни) ядра и асоциативни . Чрез превключващите ядра на таламуса се предават аферентни влияния от всички рецептори в тялото. Асоциативните ядра получават импулси от превключващите ядра и осигуряват тяхното взаимодействие. В допълнение към тези ядра, таламусът съдържа неспецифични ядра, които имат както активиращи, така и инхибиращи ефекти върху малки участъци от кората.
Благодарение на широките връзки, таламусът играе решаваща роля в живота на тялото. Импулсите, идващи от таламуса към кората, променят състоянието на кортикалните неврони и регулират ритъм на кортикална активност . С прякото участие на таламуса възниква формирането на условни рефлекси и развитието на двигателните умения, формирането на емоции човек, неговите изражения на лицето. По-специално таламусът играе важна роля в генерирането на усещания усещания за болка . Свързан с работата му регулиране на биоритмите в човешкия живот (ежедневни, сезонни и т.н.).
Хипоталамус е най-висшият подкорков център на регулация автономни функции , заявява будност и сън . Тук са вегетативните центрове, регулиране на метаболизма и тялото, предоставяйки поддържане на постоянна телесна температура (при топлокръвни) и нормално ниво кръвно налягане поддържащ воден баланс, регулиране чувство на глад и ситост . Дразненето на задните ядра на хипоталамуса предизвиква засилване на симпатиковите влияния, а на предните - парасимпатиковите ефекти.
Поради връзката на хипоталамуса с хипофизната жлеза ( хипоталамо-хипофизна система) дейността на ендокринните жлези се контролира. Вегетативните и хормоналните реакции, регулирани от хипоталамуса, са компоненти на човешките емоционални и двигателни реакции.
Неспецифична мозъчна система
Неспецифична системазаема средната част на мозъчния ствол. Не е свързано с анализа на някаква специфична чувствителност или с изпълнението на определени рефлексни реакции. Импулсите влизат в тази система чрез странични разклонения от всички специфични пътища, в резултат на което се осигурява тяхното широко взаимодействие. За неспецифична системаместоположението на невроните под формата на дифузна мрежа, изобилието и разнообразието на техните процеси са характерни. Поради тази причина тя получи името ретикуларна формация или ретикуларна формация.
Разграничете два вида влияние неспецифична система за работата на други нервни центрове активиращи и инхибиращи . И двата вида тези въздействия могат да бъдат възходящи (към лежащите центрове) и низходящи (към долните центрове). Служат за регулиране на функционалното състояние на мозъка, нивото на будност и регулиране на постурално-тоничните и фазовите реакции на скелетните мускули.
Малък мозък
Малкият мозък е надсегментно образувание, което няма пряка връзка с изпълнителния апарат.Малкият мозък се състои от нечифтно образувание - вермис и сдвоени полукълба.
Основните неврони на кората на малкия мозък са многобройни клетки на Пуркиние. Благодарение на широките връзки (до 200 000 синапса завършват във всяка клетка), интегриране на голямо разнообразие от сензорни въздействия , предимно проприоцептивни, тактилни и вестибуларни. Представителството на различни периферни рецептори в кората на малкия мозък има соматотопна организация (гръцки somatos тяло, топос място), т.е. отразява реда на тяхното местоположение в човешкото тяло. В допълнение, това разположение съответства на същото разположение на представителството на частите на тялото в мозъчната кора, което улеснява обмена на информация между кората и малкия мозък и осигурява тяхната съвместна дейност при контролиране на човешкото поведение. Правилната геометрична организация на церебеларните неврони определя нейната значение при обратното броене и точно поддържане на темпото на цикличните движения.
Основната функция на малкия мозък е регулирането на постуралните тонични реакции и координацията на двигателната активност.(Орбели Л.А., 1926 г.).
от анатомични особености(връзки на кората на малкия мозък с неговите ядра) и функционалното значение на малкия мозък се разделя на три надлъжни зони:
- вътрешна или средна кора на червея, чиято функция е регулиране на тонуса на скелетната мускулатура, поддържане на стойката и баланса на тялото;
- междинен средна частмозъчната корамалкия мозък, чиято функция е да координира постуралните реакции с движенията и да коригира грешките;
- латерална или странична мозъчна корамалък мозък, който заедно с диенцефалона кората на главния мозък участва в програмирането на бързи балистични движения (хвърляния, удари, скокове и др.).
Базални ядра
Базалните ганглии включват стриатума, състоящ се от опашното ядро и черупката, и бледото ядро и понастоящем също се считат амигдала(отнасящи се до вегетативни центровелимбична система) и substantia nigra на междинния мозък.
Аферентните влияния идват към базалните ядра от рецепторите на тялото през таламуса и от всички области на мозъчната кора. Те навлизат почти изключително в стриатума. Еферентните влияния от него се изпращат до бледото ядро и по-нататък до стволовите центрове на екстрапирамидната система, а също и през таламуса обратно към кората.
Базалните ядра участват в образуването на условни рефлекси и изпълнението на комплекса безусловни рефлекси(отбранителни, хранителни и др.). Те осигуряват необходимото положение на тялото по време на физическа работа, както и протичането на автоматични ритмични движения (древни автоматизми).
Бледото ядро изпълнява основната двигателна функция, а стриатумът регулира неговата дейност. Понастоящем е разкрито значението на опашното ядро в контрола на сложни психични процеси - внимание, памет, откриване на грешки.
Всички функции на тялото могат условно да бъдат разделени на соматични, или животно(животни), свързани с възприятието външна информацияи дейността на мускулите, и вегетативна (вегетативна), свързана с дейността на вътрешните органи, процесите на дишане, кръвообращение, храносмилане, отделяне, метаболизъм, растеж и възпроизводство.
Функционална организация на вегетативната нервна система
автономна нервна система наречен набор от еферентни нервни клетки на гръбначния и главния мозък, както и клетки на специални възли (ганглии), които инервират вътрешните органи. Дразненията на различни рецептори на тялото могат да причинят промени както в соматичните, така и в автономните функции, тъй като аферентните и централните участъци на тези рефлексни дъги са общи. Те се различават само по еферентните си дялове. характерна особеностеферентните пътища, включени в рефлексните дъги на вегетативните рефлекси, е тяхната двуневронна структура(единият неврон е в централната нервна система, другият е в ганглиите или в инервирания орган).
Вегетативна нервна системаразделена на два отдела симпатикова и парасимпатикова (фиг. 4).
Еферентни пътища симпатикова нервна системазапочва в гърдите лумбални областигръбначен мозък от невроните на страничните му рога. Прехвърлянето на възбуждане от предвъзловите симпатикови влакна към постнодалните става с участието на медиатора ацетилхолин , и от постнодални влакна до инервирани органи с участието на медиатор норепинефрин . Изключение правят влакната, които инервират потните жлези и разширяват съдовете на скелетните мускули, където възбуждането се предава с помощта на ацетилхолин.
Еферентни пътища парасимпатикова нервна системазапочват в мозъка от някои ядра на средата и продълговатия мозък, а в гръбначния мозък от неврони сакрален отдел. Провеждането на възбуждане в синапсите на парасимпатиковия път става с участието на медиатора ацетилхолин . Вторият еферентен неврон е разположен в или близо до инервирания орган.
Най-висшият регулатор на автономните функции е хипоталамусът. , който действа съвместно с ретикуларната формация и лимбичната система под контрола на мозъчната кора. Освен това невроните, разположени в самите органи или в симпатикови възли, могат да извършват собствени рефлексни реакции без участието на централната нервна система "периферни рефлекси" .
Ориз. 4. Автономна нервна система
Изходна зона на ляво влакно: парасимпатиково (черно)
и симпатични (засенчени) системи.
Вдясно структурата на еферентната част на рефлексната дъга на вегетативните рефлекси. Вляво е диаграма на средния, медулата и гръбначния мозък.
Арабски цифри номера на гръдни сегменти, римски номера на лумбални сегменти.
Функции на симпатиковата нервна система
С участието на симпатиковата нервна система възникват много важни рефлекси в тялото, насочени към осигуряване на неговото активно състояние, включително неговата двигателна активност.Те включват рефлекси разширяване на бронхите, повишена и повишена сърдечна честота , разширяване на съдовете на сърцето и белите дробове с едновременно стесняване на съдовете на кожата и органите коремна кухина (осигуряване на преразпределението на кръвта ), освобождаването на отложената кръв от черния дроб и далака, разграждането на гликогена до глюкоза в черния дроб ( мобилизиране на въглехидратни енергийни източници ), повишена активност на жлезите с вътрешна секреция и потните жлези. Симпатикова нервна система намалява активността на редица вътрешни органи: в резултат на вазоконстрикция в бъбреците, процесите на уриниране намаляват, секреторната и двигателната активност на органите се инхибира стомашно-чревния тракт; актът на уриниране е възпрепятстван мускулите на стената се отпускат Пикочен мехура сфинктерът му е намален.
Повишената активност на тялото е придружена от симпатична рефлекс на разширяване на зеницата . От голямо значение за двигателната активност на тялото е трофично влияние на симпатиковите нерви върху скелетните мускули , подобряване на техния метаболизъм и функционално състояние, облекчаване на умората.
Симпатиковият отдел на нервната система не само повишава нивото на функциониране на тялото, но и мобилизира своите скрити функционални резерви , активира мозъчната дейност, повишава защитните реакции ( имунни реакции, бариерни механизмии др.), предизвиква хормонални реакции. Симпатиковата нервна система е от особено значение. в развитието стресови състояния , в най трудни условияжизненоважна дейност. Л. А. Орбели подчерта съществено значениесимпатични влияния за адаптация (адаптация) на организма към тежка работа, към различни условиявъншна среда. Тази функция беше наименувана адаптивно-трофичен.
Функции на парасимпатиковата нервна система
парасимпатикова нервна системаизвършва стесняване на бронхите, забавяне и отслабване на сърдечните контракции: свиване на съдовете на сърцето; попълване на енергийните ресурси (синтез на гликоген в черния дроб и засилване на храносмилателните процеси); укрепване на процесите на уриниране в бъбрецитеи осигуряване на акта на уриниране (свиване на мускулите на пикочния мехур и отпускане на неговия сфинктер) и др. Парасимпатиковата нервна система осигурява главно стартови влияния : свиване на зеницата, бронхите, активиране на дейността храносмилателни жлезии така нататък.
Дейността на парасимпатиковия отдел на автономната нервна система е насочена към тока регулиране на функционалното състояние, за поддържане на постоянство вътрешна средахомеостаза . Парасимпатиковият дял осигурява възстановяване на различни физиологични параметри , драстично променена след напрегнати мускулна работапопълване на изразходваните енергийни ресурси. Посредник парасимпатикова системаацетилхолин, намалявайки чувствителността на адренергичните рецептори към действието на адреналин и норепинефрин, има определен антистрес ефект .
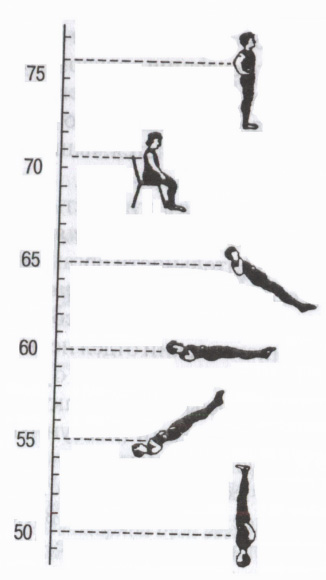
Ориз. 5. Вегетативни рефлекси
Ефект на позицията на тялото върху сърдечната честота (bpm). (Според: Могендович М.Р., 1972 г.)
Чрез автономните симпатикови и парасимпатикови пътища централната нервна система осъществява някои автономни рефлекси, като се започне от различни рецептори на външната и вътрешната среда: висцеро-висцерални (от вътрешни органи към вътрешни органи - например дихателно-сърдечен рефлекс); дермо-висцерален (от кожата - промяна в дейността на вътрешните органи по време на дразнене активни точкикожа, като акупунктура, акупресура); от рецепторите на очната ябълка око-сърдечен рефлекс на Ашнер (намаляване на сърдечната честота при натискане на очни ябълкипарасимпатиков ефект); моторно-висцерални - например, ортостатичен тест(ускоряване на сърдечната честота при преминаване от легнало в изправено положение – симпатиков ефект) и др. (фиг. 5). Те се използват за оценка на функционалното състояние на организма и особено на състоянието на вегетативната нервна система (оценка на влиянието на нейния симпатиков или парасимпатиков отдел).
лимбична система
Под лимбична системаразбират редица корови и подкорови структури, чиито функции са свързани с организацията мотивационно-емоционални реакции, памет и процеси на учене .
Кортикалните участъци на лимбичната система, представляващи нейната по-висока част, са разположени на долната и вътрешната повърхност на мозъчните полукълба (участъци на предната кора, cingulate gyrus или лимбична кора, хипокампус и др.). ДА СЕ подкорови структуриЛимбичната система включва хипоталамуса, някои ядра на таламуса, средния мозък и ретикуларната формация. Между всички тези отдели има тесни директни и обратни връзки, образуващи така наречения "лимбичен пръстен".
Лимбичната система участва в голямо разнообразие от прояви на активността на тялото в регулирането на поведението при хранене и пиене, цикъла сън-събуждане, в процесите на формиране на следа от паметта (отлагане и извличане от паметта), в развитието на агресивно-защитни реакции. осигуряване на избирателно поведение. Тя създава положителни и негативни емоции с всички двигателни, вегетативни и хормонални компоненти. Електрическата стимулация на различни части на лимбичната система чрез имплантирани електроди (при експерименти с животни, в клиниката при лечение на пациенти) разкрива наличието на центрове за удоволствиекоито генерират положителни емоции и недоволствокоито предизвикват негативни емоции. Изолираното дразнене на такива точки в дълбоките структури на човешкия мозък предизвикваше появата на чувство на „безпричинна радост“, „безсмислен копнеж“, „необясним страх“.
Функции на кората на главния мозък
При висшите бозайници и човека водещият отдел на ЦНС е кората на главния мозък.
Кортикални неврони
Кортексът е слой от сиво вещество с дебелина 2-3 mm, съдържащ средно около 14 милиарда нервни клетки. Характерно за него е изобилието от междуневронни връзки, чийто растеж продължава до 18 години, а в някои случаи и по-нататък.
Основните видове кортикални клетки са пирамидални и звездовидни неврони. звездовидни невронисвързани с процесите на възприемане на стимули и обединяването на дейностите на различни пирамидални неврони.
Пирамидални неврони изпълняват еферентната функция на кората (главно чрез пирамидалния тракт) и viutricorcal процеси на взаимодействие между отдалечени неврони. Най-големите пирамидални клетки, гигантските пирамиди на Бетц, са разположени в предния централен гирус (моторна кора).
Функционалната единица на кората евертикална колонавзаимосвързани неврони. Вертикално удължени големи пирамидални клетки с неврони, разположени над и под тях, образуват функционални асоциации на неврони. Теглото на невроните на вертикалната колона отговаря на една и съща аферентна стимулация (от същия рецептор) със същата реакция и съвместно образува еферентни отговори на пирамидални неврони. При необходимост вертикалните колони могат да се комбинират в по-големи образувания, осигурявайки сложни реакции.
Функционално значение на различни кортикални полета
Според структурните особености и функционалното значение на отделните корови области цялата кора се подразделя на три основни групи първични, вторични и третични полета (фиг. 6).
Първични полетасвързани със сетивните органи и органите за движение по периферията. Те осигуряват появата на усещания . Те включват например областта на болката и мускулно-ставната чувствителност в задната централна извивка на кората, зрителното поле в тилната област, слуховото поле във временната област и двигателното поле в предната централна извивка. В първичните полета са високоспециализирани детерминантни клетки или детектори, селективно реагиращи само на определени стимули. Например в зрителния кортекс има детекторни неврони, които се възбуждат само когато светлината е включена или изключена, чувствителни само към нейния определен интензитет, към определени интервали на излагане на светлина, към определена дължина на вълната и т.н. С разрушаването на първичните полета на кората възниква т. нар. корова слепота, корова глухота и др.
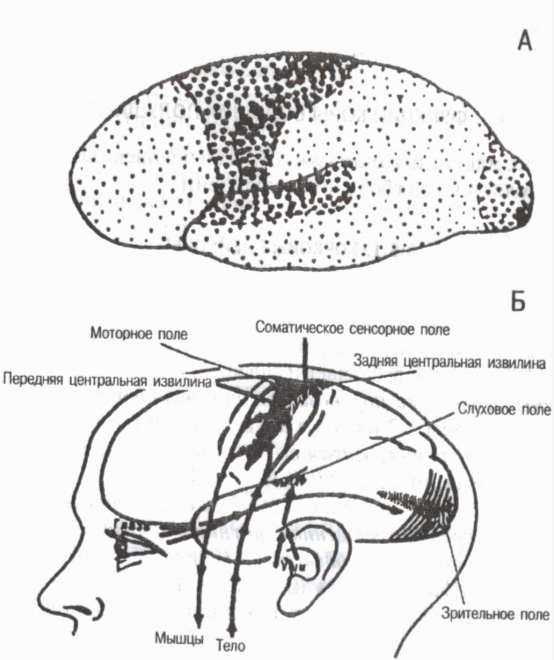
Ориз. 6. Първични, вторични и третични полета на кората на главния мозък
На А: големи точки първични полета, средни вторични полета, малки третични полета (според G. I. Polyakov 1964, A. R. Luria, 1971)
На B: първични (проекционни) полета на мозъчната кора (според W. Penfield, L. Robert, 1964)
Вторични полетаразположен до първичния. В тях се случва разбиране и разпознаване на звукови, светлинни и други сигнали възникват сложни форми на обобщено възприятие . При засягане на вторичните полета се запазва способността да се виждат предмети, да се чуват звуци, но човекът не ги разпознава, не помни смисъла.
Третични полетаразвит почти изключително при хора. Това са асоциативни области на кората, които осигуряват най-високите форми на анализ и синтез и формират целенасочената поведенческа активност на човек. Третичните полета са разположени в задната половина на кората между теменната, тилната и темпоралната област и в предната половина в предните части фронтални области. Тяхната роля е особено важна в организацията на координираната работа на двете полукълба . Третичните полета узряват при хората по-късно от други кортикални полета и се разграждат по-рано от други по време на стареенето.
Функцията на задните третични полета (главно долните париетални области на кората) е приемането, обработката и съхранението на информация. Те формират представата за схема на тялото и схема на пространството , осигуряващи пространствена ориентация на движенията. Предни третични полета (антерофронтални области) извършва обща регулация на сложни форми на човешкото поведение, формиране на намерения и планове, програми за произволни движения и контрол върху тяхното изпълнение . Развитието на третичните полета при хората е свързано с говорна функция. мислене ( вътрешна реч) е възможно само при съвместната дейност на различни сензорни системи, обединяването на информацията от които се извършва в третичните полета. При вродено недоразвитие на третичните полета човек не е в състояние да овладее речта (произнася само безсмислени звуци) и дори най-простите двигателни умения (не може да се облича, да използва инструменти и др.).
Двойна активност и доминиране на полукълбата
В резултат на това се извършва обработка на информация сдвоена активност на двете полукълба мозък. Въпреки това, като правило, едно от полукълбата е водещо доминантно . Повечето хора с водещи дясна ръка(десняк) лявото полукълбо е доминиращо, а дясното полукълбо е подчинено (субдоминантно).
Ляво полукълбов сравнение с дясно има по-фина невронна структура, по-голямо богатство на невронни връзки, по-концентрирано представяне на функции и По-добри условиякръвоснабдяване. В лявото доминантно полукълбо е моторният център на речта (центърът на Брока), който осигурява речева дейности сензорния център на речта, който разбира думите. Лявото полукълбо е специализирано във финия сензомоторен контрол на движенията на ръцете.
Хората имат три форми функционална асиметрия: моторни, сензорни и умствени . По правило човек има водеща ръка, крак, око и ухо. Проблемът с функционалната асиметрия обаче е доста сложен. Например, човек с дясна ръка може да има доминиращо ляво око или лявото ухо, сигналите от които са доминиращи. В същото време във всяко полукълбо могат да бъдат представени функциите не само на противоположната, но и на същата страна на тялото. В резултат на това е възможно да се замени едно полукълбо с друго в случай на повреда, а също и създава структурна основа за променлива хемисферична доминация в моторния контрол.
Психическата асиметрия се проявява под формата на определен полукълбо специализации. За ляво полукълбо Характерни са аналитични процеси, последователна обработка на информация, включително с помощта на реч, абстрактно мислене, оценка на времеви връзки, предвиждане на бъдещи събития и успешно решаване на вербално-логически проблеми. IN дясно полукълбо информацията се обработва холистично, синтетично (без да се разделя на детайли), като се отчита минал опит и без участието на речта, преобладава обективното мислене. Тези характеристики позволяват да се свърже с дясното полукълбо възприемането на пространствени характеристики и решаването на зрително-пространствени задачи. Функциите на дясното полукълбо са свързани с миналото, а на лявото с бъдещето.
Електрическа активност на кората на главния мозък
Промените във функционалното състояние на кората се отразяват в неговия запис електрическа активност електроенцефалограма (ЕЕГ). Съвременните електроенцефалографи усилват потенциала на мозъка 2-3 милиона пъти и дават възможност за изследване на ЕЕГ от много точки на кората едновременно, т.е. системни процеси. EEG регистрациянаправени под формата на мастило върху хартия, както и във формата пълна картинавърху повърхността на мозъка, т.е. карти на мозъка (метод на картографиране) върху екрана на монитора на съвременни компютърни системи (фиг. 7).
Има определени честотни диапазони , Наречен ЕЕГ ритми (фиг. 8): в състояние на относителна почивка най-често се записва алфа ритъмът (8-13 трептения за 1 s); в състояние на активно внимание - бета-ритъм (14 трептения в 1s и повече); при заспиване някои емоционални състояниябета ритъм (4-7 трептения в 1s); с дълбок сън, загуба на съзнание, анестезия-делта ритъм (1-3 флуктуации за 1 s).
ЕЕГ отразява характеристиките на взаимодействието на кортикалните неврони по време на умствено и физическа работа (Ливанов M.N., 1972). Липсата на координация при извършване на необичайна или тежка работа води до т.нар Десинхронизация на ЕЕГбърза асинхронна дейност. Когато се формира двигателно умение, се появяват феномени в ЕЕГ ЕЕГ синхронизацияукрепване на взаимосвързаността (синхронност и синфазност) на електрическата активност на различни области на кората, участващи в контрола на движенията. При циклична работа се появяват бавни потенциали в темпото на извършеното, въображаемо или предстоящо движение "маркирани ритми"(Sologub E. B., 1973).
В допълнение към фоновата активност в ЕЕГ има отделни потенциали, свързани със събития:предизвикани потенциаливъзникващи в отговор на външни стимули (слухови, зрителни и др.); потенциали, отразяващи мозъчните процеси по време на подготовката, изпълнението и завършването на отделните двигателни действия това "вълна от очакване"или условна отрицателна вълна (Walter G., 1966), премоторни, двигателни и крайни потенциали и др. В допълнение, различни супер бавни трептенияс продължителност от няколко секунди до десетки минути (по-специално, така наречените "омега-потенциали" и др.), които отразяват биохимичните процеси на регулиране на функциите и умствената дейност.

Ориз. 7. Мозъчно картографиране
Многоканален запис на човешка електроенцефалограма (ЕЕГ) на екрана на монитора и отразяване на възбудени (светли зони) и инхибирани (тъмни зони) области на кората.
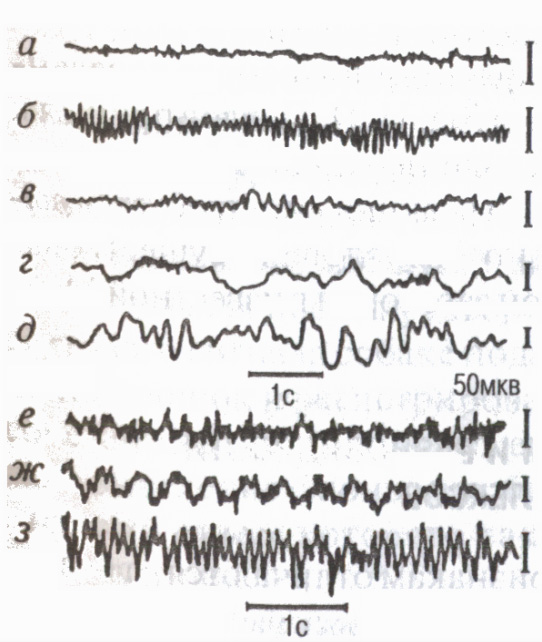
Ориз. 8. ЕЕГ на тилната (a-e) и моторната (f-h) области на кората на главния мозък на човека в различни състоянияи по време на мускулна работа
активно състояние,
отворени очи (бета ритъм); b- почивка, затворени очи (алфа ритъм);
при сънливост (теша-ритъм);
d заспиване; д дълбок сън„З
(делта ритъм); f необичайно или
тежка работа асинхронен чести
активност (феномен на десинхронизация);
g циклична работа бавни потенциали в темпото на движенията ("белязани ритми" ЕЕГ); h изпълнение на овладяното движение появата на алфа ритъма.
Медицина и ветеринария
пирамидални неврони. големи клетки, перикариони с размери 10-130 микрона. Клетката има апикален дендрит, който е насочен към молекулния слой; странични процеси - дендрити; дълъг аксон отива от основата - това ще бъде моторно нервно влакно (началото на образуването на пирамидалния тракт).
Лекция по хистология №13
Централна нервна система. Кората на главния мозък. Цитоархитектоника на слоевете на кората на главния мозък. нервен състав. Характеристика на пирамидални неврони. Модулна структурна и функционална единица на кората на главния мозък. Малък мозък. Структурата и невроналният състав на кората на малкия мозък.
Източник на развитие на сивото и бялото вещество на мозъка
Развива се от мантията или мантийната зона на сивото вещество на невралната тръба
маргинален воал бяло вещество
Композитни компонентисиво вещество на мозъка (неокортекс)
6 слоя:
- Молекулярният се формира главно от процесите на клетките на подлежащите слоеве, но има единични неврони, които се наричат хоризонтални.
- Външно гранулирано присъствиезвездовидна неврони и не многобройни пирамидални неврони.
- Външен пирамидален (пирамидален) най-широкият слой съдържапирамидални неврони.
- Вътрешна гранулирана финазвездовидна неврони, осъществяват вътрекортикални връзки (не навлизат в бялото вещество).
- Вътрешен пирамидален (ганглиозен) представен от големипирамидален неврони или клетки на Бетс.
- Полиморфните клетъчни форми са разнообразни, клетките са малки, аксоните им се простират в бялото вещество.
звездовидни неврони.Перикарионите са с кръгла или триъгълна форма, 4-8 µm в диаметър; аксонът и дендритите на тези клетки участват в интракортикалните връзки.
пирамидални неврони.Големи клетки, перикариони с размери 10-130 микрона. Клетката има апикален дендрит, който е насочен към молекулния слой; странични процеси дендрити; от основата идва дълъг аксон ще бъде двигателно нервно влакно (началото на образуването на пирамидалния тракт). Колатералите идват от аксона: те отиват към съседните клетки или се издигат и взаимодействат с горните слоеве.
основни характеристикикортикални неврони:
Всички неврони в мозъчната кора са мултиполярни. По функция, 1-4 слоя: асоциативен (вмъкване), 5-6 моторни.
Структурна и функционална единица на мозъка: модул
Модул (в мозъчната кора до 3 милиона) вертикална колона с диаметър 300 микрона, перпендикулярна на повърхността на кората и преминаваща през цялата дебелина на кората.
В центъра на модула:
- Едно кортико-картично нервно влакно е нервно влакно в кората; идващи или от пирамидални неврони на същото полукълбо (асоциативно-интеркаларно), или от противоположното полукълбо (комиссурално).
- Две таламокортикални влакна (аферентно-сензорни влакна), идващи от таламуса и преминаващи в 4 слоя; след това се образува еферентно нервно влакно образувани от аксониНай-добрите клетки.
Цитоархитектоникаучастъци от кората, които се различават един от друг по отношение на структурата и разположението на клетките.
Миелоархитектоникаучастъци от кората, които се различават един от друг по характеристиките на местоположението на влакната.
полета различни области на кората, които се различават една от друга: цито, миелоархитектоника и функционално значение.
Видове кора:
- Гранулиран чувствителен център, в който 2-ри и 4-ти слой са добре развити; те завършват с (аферентни) сензорни нервни влакна, идващи от органа на обонянието, слуха и зрението.
- Агрануларен двигателен център, в който са развити 3-ти, 5-ти и 6-ти слой.
Достоверен морфологичен еквивалент на интелекта е броят на синапсите на асоциативните неврони (слоеве 1-4), обикновено 10 хиляди.
Малък мозък
Източник на развитие на задния мозък на малкия мозък
Функции: координира движението и баланса
Малкият мозък има голям брой извивки, върху препарата под формата на разклонено дърво.
Гирусите са разделени от бразди. Всяка извивка съдържа: тясна пластина от бяло вещество, напълно покрита сива материя(кора). В дълбините на бялото вещество на малкия мозък лежат клъстери от неврони: ядра на малкия мозък.
Има три слоя в сивото вещество или кората на малкия мозък:
- Външната молекула е представена от мултиполярни асоциативни (интеркаларни) неврони. Има 2 вида клетки:
- Кошничните неврони са разположени в долната трета на молекулярния слой, клетките неправилна формамалки размери. Аксонът на клетката влиза в синапс с перикареона на клетката на Пуркение, където се разклонява и образува плексус под формата на кошничка. Дендрити в молекулярния слой.
- Звездовидните (големи и малки) са локализирани от външната страна на молекулярния слой (повърхността на кората).Големи звездовидни неврони:аксонът синапсира с перикариона на клетките на Пуркение и участва в образуването на кошничката. Дендрити в молекулярния слой.малки звездовидни неврони: Аксонът синапсира с дендритите на клетките на Пуркение.
- Среден ганглиозен - представен от един слой, тела на мултиполярни моторни неврони или клетки на Пуркиние. Това са големи крушовидни клетки от перикариона (тялото), 2-3 дендрита се простират в молекулярния слой. Аксонът напуска тялото на клетката и преминава през гранулирания слой в бялото вещество, където завършва при ядрата на малкия мозък. Аксоните на клетките на Пуркиние са единствените еферентни (моторни) нервни влакна, излизащи от кората на малкия мозък. По-близо до клетъчното тяло, колатералите се отклоняват от аксона, които отиват до ганглиозния слой и дълбоките части на молекулярния слой, където влизат в синапс с тела и дендритисъседни клетки на Пуркиние.
- Вътрешно гранулираните всички клетки са полярни и асоциативни съдържат два вида клетки:
1-Клетки-Зърна малки клетки, перикариони имат диаметър 4-6 микрона, дендрити: къси 3-4 парчета, насочени към бялото вещество, напомнящи на птичи крака. Аксонът се изкачва в молекулярния слой, където се разделя в Т-образна форма.
2-Голяма звездовидна: с дълги аксони и къси аксони Голджи клетки от втори тип. Големите клетки на тялото на които са локализирани непосредствено под ганглийния слой. Аксонът влиза в синапса с розетки от мъхови влакна, преди те да влязат в синапса с дендрити на гранулирани клетки. Дендритите се издигат в молекулярния слой.
Аферентни (сензорни) нервни влакна:
- Бриофитни влакна Краен бриофит нервно влакносе нарича гнездо. Розетки на синапса на мъхестите влакна с дендритите на зърнестите клетки, образуващи мозъчни гломерули.
- Катерещи или лианоподобни влакна идват от бялото вещество, преминават през гранулираните слоеве и влизат в синапс или с перикариона на клетката на Пуркиние, или с дендритите на тези клетки.
Аксоните на зърнестите клетки влизат в синапса с дендрити:
- клетъчен молекулен слой
- Cell Purkenye
- Голджи клетки от втори тип
Инхибиторната система на малкия мозък включва:
- Клетки от молекулярния слой
- Голджи клетки от втори тип
- Възбудни синапси:
- Мъхови влакна
- Лианоидни влакна
Както и други произведения, които може да ви заинтересуват |
|||
| 73026. | Въвеждане, форматиране на данни и писане на формули | 126KB | |
| Цел на работа: С помощта на командата Format Cell форматирайте данните в колона D с паричен формат без десетични знаци. С помощта на бутона Format Painter копирайте формата на колона D в E. Използвайте бутоните на лентата за форматиране, за да зададете колона F в процентен формат с два знака след десетичната запетая. | |||
| 73027. | Изграждане и редактиране на диаграми в Excel | 160KB | |
| Научете как да създавате диаграми с помощта на съветника; Научете как да редактирате диаграми. Контролни въпроси Какво е предназначението на диаграмите Какви видове диаграми познавате Как се изгражда диаграма отделен листКак да промените типа на диаграмата Как да изтриете диаграма... | |||
| 73028. | Моделиране на файлови системи | 147,5 КБ | |
| Потребителите дават на файловете символични имена, като се вземат предвид определени ограничения на операционната система. Директорията съдържа списък с включени в нея файлове и се установява съответствие между файловете и техните характеристики и атрибути. | |||
| 73029. | Обозначаването на структурно-фазовия склад на HBM, който трябва да компенсира VNT, чрез методите на рентгенова дифракция и електронна микроскопия | 1,5 MB | |
| Определете структурно-фазовата структура на HBM, за да отмъстите на VNT за данни от рентгенова дифракция и електронна микроскопия. Проследяване на промяната в структурно-фазовия склад на НВМ в процеса на термохимична обработка. | |||
| 73033. | Персонализирани типове данни на примера на MS SQL Server | 51KB | |
| Да се проучат възможностите за дефиниране и използване на потребителски типове данни, DML и DDL тригери, ограничения за интегритет в пост-релационна СУБД. Овладейте методите и технологиите за създаване на потребителски типове данни в CLR.Net. | |||
| 73034. | Принципът на маркиране на стоманобетонни подови плочи на граждански сгради | 47,1 КБ | |
| Принципът на маркиране на стоманобетонни подови плочи на граждански сгради В съответствие с GOST 2300978 подовите плочи са маркирани с редица буквено-цифрови индекси. Първата група съдържа обозначението на типа на плочата и нейните конструктивни размери, дължина и ширина в дециметри. | |||
Нервна система. Лекция 2
Кораголям мозък (cortex cerebri) представлява непрекъснат слой сиво вещество върху повърхността на мозъчните полукълба с дебелина 2-5 mm. Той е разположен не само върху извивките, но продължава във всички бразди, поради което площта му е сравнително голяма и възлиза на 2200 cm 2 при възрастен. Масата на кората е 581 g, обемът й е около 560 cm3. В същото време основният обем на кората е съставен от бяло вещество (450 cm 3, тегло - 470 g). Телесното тегло на кортикалните неврони е само около 21 g (20 cm 3).
Размерът на кортикалните нервни клетки варира в широки граници от 8-9 мкдо 150 мк.В мозъчната кора на човека има до 15 милиарда нервни клетки, от които 6 милиарда са малки клетки. По-голямата част от кортикалните неврони са два типа: пирамидални неврони и звездовидни неврони. Тези клетки са разположени в кората на определени слоеве.
В човешкия мозък филогенетично има няколко различни видовекора:
Древната кора - палеокортекс (0,6%) - наслояване практически липсва (1-2 слоя), локализирано в областта на обонятелните триъгълници, образува покритие на прозрачната преграда и обгражда ядрото на амигдалата.
Старата кора - архикортекс (2,2%) - има 2-3 слоя, представена е от сивото вещество на зъбчатия гирус, сивото вещество на хипокампуса и мозъчните ивици на corpus callosum.
Преходна кора - мезокортекс (1,6%) - област на сводестия гирус.
Новият кортекс, неокортексът (95,6%), е добре структуриран и има 6 слоя.
При висшите гръбначни животни има предимно шест повече или по-малко ясно определени слоя. Но всеки от тези слоеве, с изключение на първия, може да бъде разделен на два или дори три подслоя.
Първи слой, така наречените зонални или молекулярна плоча, се състои главно от плексуси от апикални дендрити на пирамидални неврони, клетъчните тела на които са разположени в други слоеве на кората. В първия слой има много малко нервни клетки. Представлява хоризонтални клетки, чиито аксони и дендрити също са разположени хоризонтално в същия слой.
Втори слой, така наречената външна гранулирана или външна гранулирана плоча, включва маса от малки клетки, принадлежащи към малки пирамидални, така наречените интеркаларни неврони, и звездовидни неврони, с малки пирамидални клетки, доминиращи във втория слой.
Третият слой е външната пирамидална плочасъдържа пирамидални клетки със среден размер
четвърти слой, така наречената вътрешна гранулирана или вътрешна гранулирана ламина, се състои главно от малки звездовидни клетки, но също така съдържа малки и средни по размер пирамиди.
Пети слой - вътрешната пирамидална плочасъдържа гигантски пирамидални клетки или клетки на Betz.
Шести слой – многоформена плочасъдържат предимно пирамидални клетки със среден размер и малко количество малки пирамидални и звездовидни клетки.
Първите три слоя са най-младите, те осигуряват връзка между различните части на кората. Четвъртият слой е максимално развит в областите, където идва аферентна информация (чувствителни центрове, особено постцентралната извивка). Петият слой се изразява в двигателните зони на кората: прецентрален гирус, перицентрален лобул, супрамаргинален извивка.
Всеки пирамидален неврон има клетка с форма на пирамида и множество дендрити. Аксонът на пирамидален неврон излиза от малка могила в основата на клетката. В малките, така наречените интеркаларни или междинни пирамидални неврони, аксоните, разклонени в хоризонтална или вертикална посока, завършват веднага, без да напускат кората. Аксоните на средни и големи пирамиди отделят много колатерали в кората, а основните стъбла отиват в подкоровото бяло вещество. Някои от тях се връщат от подкоровото вещество в кората на дадено полукълбо или, преминавайки през corpus callosum, завършват в кората на другото полукълбо. Те служат за обединяване на различни части на мозъчната кора. Следователно такива пирамидални неврони се наричат асоциативен.Други аксони отиват в подкорови образувания и по-нататък в различни части на мозъка и гръбначния мозък. Тези пирамиди се наричат проекция.Във връзка с филогенетичното развитие на бозайниците броят на пирамидните клетки се увеличава значително.
Широчината на цялата кора, броят на клетките и ширината на всеки слой от клетки в предната част, както и клетъчният състав, т.е. размерът, формата и разположението на клетките във всяка секция на кората, варират значително. Тези вариации заемат определени, ясно обособени територии. В големия мозък на бозайниците има 11 добре дефинирани големи кортикални полета.
Въз основа на хистологични и физиологични изследвания всяко такова поле беше разделено на няколко по-независими секции. По-значителна диференциация на кората се наблюдава при висшите бозайници - маймуните. Особено голямо е при хората.
Началото на такива изследвания е положено от киевския учен Владимир Алексеевич Бетс, който през 1874 г. публикува статия „Два центъра в мозъчната кора“, в която описва двигателната зона в прецентралната извивка и сензорната зона в постцентралната.
През 1909 г. немският невролог Корбиниан Бродман публикува карти на цитоархитектоничните полета. мозъчната кора. Бродман е първият, който създава карти на земната кора. Впоследствие O. Vogt и C. Vogt (1919-1920), като вземат предвид структурата на влакната, описват 150 миелоархитектонични области в кората на главния мозък. В Института по мозъка на Академията на медицинските науки на СССР И. Н. Филипов и С. А. Саркисов създават карти на кората на главния мозък, включващи 47 цитоархитектонични полета.
Данни експериментални изследванияпоказват, че разрушаването или отстраняването на определени области на мозъчната кора при животни нарушава определени функции. Тези факти се потвърждават клинични наблюденияза болни хора с туморни лезии или наранявания на някои части на мозъчната кора. Всичко това доведе до заключението, че в кората на главния мозък има центрове, които регулират изпълнението на определени функции. Морфологично потвърждение на данните от физиологията и клиниката беше доктрината за разнородната структура на кората на мозъчните полукълба в различните му части - цито- и миелоархитектониката на кората. Установено е, че невроните са разположени в кората не дифузно, а са групирани в ансамбли.
Използването на съвременни микроелектродни методи за изследване на функциите на кортикалните неврони значително разшири разбирането за обработката на сензорна информация в неокортекса и структурната организация на кората. През 1957 г. американският изследовател У. Маунткасъл, анализирайки реакциите на клетките в соматосензорната (сензорна моторна) кора на котка към стимули от различни модалности, открива следното интересен факт. Когато микроелектродът беше потопен перпендикулярно на повърхността на соматосензорния кортекс, всички клетки, които срещна, реагираха по един и същи начин на стимул, например на леко докосване на кожата или на движение в ставата. Ако електродът е потопен под ъгъл към повърхността на кората, тогава по пътя му се срещат неврони с различна сензорна модалност, редуващи се с определена периодичност.
Въз основа на тези експериментални факти У. Маунткасъл стига до извода, че соматосензорната кора е организирана в елементарни функционални единици - колониориентирани перпендикулярно на повърхността. Диаметърът на такава колона, около 500 µm, се определя от хоризонталното разпределение на терминалите на аферентното таламокортикално влакно и вертикалната ориентация на дендритите. пирамидални клетки. Според Маунткасъл колоната е елементарен блок на сензомоторната кора, където се извършва локална обработка на информация от рецептори от същата модалност. Тази хипотеза за колонната организация на неокортекса беше широко приета и даде тласък на по-нататъшни изследвания в тази област у нас и в чужбина.
Според модерни идеи, всяка функционална колона на сензомоторната кора се състои от няколко морфологични микромодула, които комбинират пет или шест вложени неврона. Съставът на такъв модул включва няколко пирамидални клетки, чиито апикални дендрити са възможно най-близо и образуват дендритен пакет; в рамките на този лъч са възможни електротонични връзки, които по всяка вероятност осигуряват синхронната работа на цялата асоциация.
В съседство с група от вертикално ориентирани пирамидални клетки са звездовидни клеткис които влизат в контакт идващите към микромодула таламокортикални влакна. Някои от инхибиторните звездовидни клетки имат дълги аксони, които се простират хоризонтално. Аксоните на пирамидалните клетки образуват повтарящи се колатерали, които могат да осигурят както улесняващи влияния в рамките на микромодула, така и инхибиторни взаимодействия между микромодулите чрез контакт с инхибиторни интерневрони. Няколко структурни микромодула, обединени чрез хоризонтално разклоняване на терминалите на специфични таламокортикални аференти, аксони на терминали и процеси на звездни клетки, образуват колона (или макромодул), чийто диаметър достига 500-1000 µm. Всяка колона се характеризира с функционално единство, което се проявява във факта, че невроните на колоната реагират на стимул от една модалност.
Впоследствие принципът на колонната организация беше потвърден в изследването на др проекционни зоникора.
И.П. Павлов разглежда кората на главния мозък като непрекъсната възприемаща повърхност, като набор от кортикални краища на анализатори. Той показа, че кортикалния край на анализаторите не е строго определена зона. В кората на главния мозък се разграничават ядро и разпръснати елементи. Ядрото е мястото на концентрация на нервните клетки на кората, които представляват точна проекция на всички елементи на определен периферен рецептор, където се извършва най-високият анализ, синтез и интеграция на функциите. Разпръснатите елементи могат да бъдат разположени както по периферията на ядрото, така и на значително разстояние от него. Те са по-прост анализ и синтез. Наличието на разпръснати елементи по време на разрушаването на ядрото отчасти позволява да се компенсира нарушената функция. Площи, заети от разпръснати елементи различни анализатори, могат да се припокриват, да се припокриват. По този начин кората на главния мозък може схематично да се представи като набор от ядра на различни анализатори, между които има разпръснати елементи, свързани с различни (съседни) анализатори. Горното ни позволява да говорим за динамична локализация на функциитев кората на главния мозък (I.P. Pavlov).
Нека разгледаме позицията на някои кортикални краища на различни анализатори (ядра) по отношение на извивките и дяловете на мозъчните полукълба при хората в съответствие с цитоархитектоничните карти.
1. В кората на постцентралния гирус и горния париетален лобул има нервни клетки, които образуват сърцевината на кортикалния анализатор на общия чувствителност(температурни, болкови, тактилни) и проприоцептивни. Провеждащите сензорни пътища, следващи мозъчната кора, се пресичат или на нивото на различни сегменти на гръбначния мозък (пътища на болка, температурна чувствителност, допир и натиск), или на нивото на продълговатия мозък (пътища на проприоцептивна чувствителност в кортикална посока). В резултат на това постцентралните извивки на всяко от полукълбата са свързани с противоположната половина на тялото. В постцентралния гирус всички рецепторни полета на различни части на човешкото тяло са проектирани по такъв начин, че кортикалните краища на анализатора на чувствителността на долните части на тялото са разположени най-високо и долни крайници, а най-ниските (по-близо до страничната бразда) са рецепторните полета на горните части на тялото и главата, горните крайници.
2. Ядро моторанализаторът е разположен главно в така наречената моторна зона на кората, която включва прецентралния гирус и парацентралния лобул на медиална повърхностполукълбо. В 5-ия слой на кората на прецентралната гирусна лъжа пирамидални неврони (клетки на Бетц),които I.P. Павлов нарича интеркаларни и отбелязва, че тези клетки с техните процеси са свързани с подкоровите ядра, двигателните клетки на ядрата на черепа и гръбначномозъчни нерви. Освен това в горните части на прецентралната извивка и в парацентралната лобула има клетки, импулсите от които са насочени към мускулите на долните части на тялото и долните крайници. В долната част на прецентралната извивка са двигателните центрове, които регулират дейността на мускулите на лицето.
По този начин всички части на човешкото тяло се проектират с главата надолу в прецентралната извивка. Поради факта, че пирамидалните пътища, произхождащи от гигантски пирамидални клетки, се пресичат или на нивото на мозъчния ствол (кортикално-ядрени влакна), на границата с гръбначния мозък, или в сегменти на гръбначния мозък (кортикално-спинален тракт), двигателните зони на всяко от полукълбата са свързани със скелетните мускули на противоположната страна на тялото. Ако мускулите на крайниците са изолирани от едно от полукълбата, мускулите на тялото, ларинкса и фаринкса са свързани с двигателните зони на двете полукълба.
Мотор I.P. Павлов също нарича областта на мозъчната кора рецепторна област, тъй като тук анализът на проприоцептивните (кинетични стимули, възприемани от рецептори, вградени в скелетни мускули, сухожилия, фасции и ставни капсули).
3. Ядрото на анализатора, осигуряващо функцията комбинирано завъртане на главата и очите навътре обратната страна , е в задни отделисредна фронтална извивка, в така наречената премоторна зона.
4. В областта на долния париетален лобул, в супрамаргиналната извивка, има ядрото на моторния анализатор, чието функционално значение е да изпълнява синтезвсичко целенасочени, професионални, трудови и спортни движения.Това ядро е асиметрично. За десничарите е в лявата, а за левичарите- само в дясното полукълбо.Способността за координиране на тези сложни целенасочени движения се придобива от индивида през живота му в резултат на практическа дейност и натрупване на опит. Изпълнението на целенасочени движения се дължи на образуването на временни връзки между клетките, разположени в прецентралната и супрамаргиналната извивка. Поражението на полето не причинява парализа, а само води до загуба на способността да се произвеждат сложни координирани целенасочени движения - апраксия (праксис - практика).
5. В кората на горната париетална лобула е сърцевината на кожния анализатор на един от специфичните видове чувствителност, който има функцията разпознаване на предмети чрез допир,- стреогнозия. Кортикалният край на този анализатор се намира в дясното полукълбо. Поражението на повърхностните слоеве на кората в този раздел е придружено от загуба на функцията за разпознаване на обекти чрез допир, въпреки че други видове обща чувствителност са запазени.
6. В дълбочината на страничния сулкус, на повърхността на средната част на горната темпорална извивка, обърната към островчето (където се вижда напречната темпорална извивка или извивката на Geschl), има ядро слуховианализатор. ДА СЕ нервни клетки, съставляващи ядрото слухов анализаторвсяко от полукълбата, пътищата от рецепторите както на лявото, така и правилната страна. В тази връзка едностранното увреждане на това ядро не води до пълна загуба на способността за възприемане на звуци. Двустранната лезия е придружена от кортикална глухота.
7. Ядро визуаленанализаторът се намира в тилен дялмозъчни полукълба в обиколката на шпорния жлеб. Ядро зрителен анализаторна дясното полукълбо е свързано чрез пътища с латералната половина на ретината на дясното око и медиалната половина на ретината на лявото око. В кората на тилната част на лявото полукълбо се проектират съответно рецепторите на латералната половина на ретината на лявото око и медиалната половина на ретината на дясното око. Що се отнася до ядрото на слуховия анализатор, само двустранно увреждане на ядрата на зрителния анализатор води до пълна кортикална слепота. Поражението на полето, което е малко по-високо от ядрото на визуалния анализатор, е придружено от загуба на зрителна памет, но не се отбелязва загуба на зрение. Още по-високо е част от кората, поражението на която е придружено от загуба на способността за навигация в непозната среда.
8. На долната повърхност на темпоралния лоб на мозъчното полукълбо, в областта на куката и отчасти в областта на хипокампуса, има ядро обонятелнианализатор. Тези области, от гледна точка на филогенезата, принадлежат към най-древните части на мозъчната кора. Обонянието и вкусът са тясно свързани, което се обяснява с близкото разположение на ядрата на обонятелния и вкусовия анализатор. Също така беше отбелязано (В. М. Бехтерев), че вкусовото възприятие се нарушава, когато кората на най-ниските части на постцентралната извивка е повредена. Ядрата на вкусовите и обонятелните анализатори на двете полукълба са свързани с рецептори от лявата и дясната страна на тялото.
Описаните кортикални краища на някои анализатори не присъстват в кората на главния мозък. само хората, но и животните.Те са специализирани във възприемането, анализа и синтеза на сигнали, идващи от външната и вътрешната среда, които, според дефиницията на И. П. Павлов, съставляват първата сигнална система на реалността.
Втора сигнална системаприсъства само при хората и се дължи на развитието на речта. Речта и заедно с нея съзнанието са филогенетично най-младите функции на мозъка. В това отношение кортикалните краища на анализаторите са най-малко локализирани. Речта и умствените функции се осъществяват с участието на цялата кора.В мозъчната кора обаче могат да се разграничат определени области, които се характеризират със строго определени речеви функции. Да, анализатори моторна реч(устни и писмени) са разположени до моторната зона на кората, по-точно в онези части на кората на предния лоб, които са в съседство с прецентралната извивка.
Анализатори визуални и слухово възприятие речеви сигнали се намират до анализаторите на зрението и слуха. Трябва да се отбележи, че говорните анализатори при десничарите са локализирани в лявото полукълбо, а при левичарите - предимно в дясното.
Помислете за позицията в кората на главния мозък на някои от анализаторите на речта.
1. Моторният анализатор на писмената реч се намира в задната част на средната фронтална извивка. Дейността на този анализатор е тясно свързана с анализатора на целенасочените движения, който осигурява формирането на заучените движения на ръката и очите, необходими за писане. При увреждане се губи способността за извършване на фини движения, обикновено извършвани под зрителен контрол и необходими за изписване на букви, думи и други знаци (аграфия).
2. Ядрото на моторния анализатор речева артикулация(речево-двигателен анализатор) се намира в задните отдели на долния фронтален извивка (център на Broca). Увреждането на кората на тази област води до двигателна афазия, т.е. загуба на способността да се произнасят думи (афазия). Тази афазия не е свързана със загуба на способността за свиване на мускулите, участващи в производството на реч. Освен това, с поражението на това поле, способността за произнасяне на звуци или пеене не се губи.
3. В централни отделиразположена е долната фронтална извивка ядрото на речевия анализатор, свързан с пеенето.Поражението му е придружено от вокална амузия - невъзможност за композиране и възпроизвеждане на музикални фрази и аграматизъм, когато се губи способността да се съставят смислени изречения от отделни думи. Речта на такива хора се състои от набор от думи, които не са свързани по семантично значение.
4. Ядро слухов говорен анализатортясно свързано с
кортикален център на слуховия анализатор и се намира в задните отдели
горна темпорална извивка, обърната на повърхността си
латерална бразда на мозъчното полукълбо. Увреждането на ядрото не нарушава слуховото възприятие на звуците като цяло, но се губи способността за разбиране на думи и реч (вербална глухота или сензорна афазия). Функцията на това ядро е, че човек не само чува и разбира речта на друг човек, но и контролира собствената си.
5. В средната трета на горната темпорална извивка се намира ядрото на кортикалния анализатор, поражението на което е придружено от появата на музикална глухота, когато музикалните фрази се възприемат като безсмислен набор от различни шумове. Този кортикален край на слуховия анализатор принадлежи към центровете на втората сигнална система, които възприемат словесното обозначаване на обекти, действия, явления, т.е. получаване на сигнали.
6. В пряка връзка с ядрото на зрителния анализатор е ядрото визуален анализатор на писмена реч(поле 39), разположен в ъгловата извивка на долния париетален лобул. Поражението на това ядро води до загуба на способността за възприемане на писмен текст, за четене (дислексия).
Фронталната кора на човека е свързана със социален принцип. Невроните му образуват двупосочни връзки с всички останали части на кората. Нарушенията се образуват при повреда социално поведение, грубост, нелоялност, намалена целенасоченост, апатия. След лоботомията - пълна липса на инициатива и социална активност.
Асиметрията на функциите на дясното и лявото полукълбо е силно изразена. Лявото полукълбо осигурява логика, абстрактно мислене, доминира по отношение на речеви функции (четене, писане, броене), сложни произволни движения. Дясно полукълбо- емоционалност, образно мислене, неречеви функции на разпознаване на сложни визуални и слухови образи, тактилни възприятия, възприятие за пространство, форма, посока, интуитивно мислене.
Подобни статии
-
Когато съпругът е против дете, как да забременеете без негово знание?
Понякога можете да забременеете по небрежност. За да не се случи това, е важно да знаете как можете да заченете дете случайно и какви средства можете да използвате, за да избегнете нежелана бременност. Също така в тази статия можете да намерите информация за...
-
Какви камъни и амулети са подходящи за Телец според хороскопа и датата на раждане Талисман на слон за Телец
Април-май Телците (21 април - 20 май) са премерени, не са суетливи и колосално продуктивни! Тяхната завидна упоритост може да доведе другите до дръжката, но те знаят точно какво правят и защо имат нужда от това. Сред положителните...
-
Ограничения за достъп до данни в роли 1c
Всички настройки на потребителските права, които ще направим в рамките на тази статия, се намират в раздел 1C 8.3 „Администриране“ - „Настройки на потребител и права“. Този алгоритъм е подобен в повечето конфигурации на ...
-
1c стартира тънък клиент вместо дебел
Платформи: 1C: Enterprise 8.3, 1C: Enterprise 8.2, 1C: Enterprise 8.1 Конфигурации: Всички конфигурации2012-11-16 21362 Те се стартират чрез указване на специални ...
-
Доказателства за известни начини за кражба на ток Как да разберете кой краде ток
Повишаването на енергийните тарифи е една от поразителните характеристики на задълбочаващата се икономическа криза. В този контекст кражбата на електроенергия и проблемите, свързани с разкриването й, са от първостепенно значение.Начини за разкриване на кражба ...
-
Характеристики на монтаж на контакти и превключватели на различни повърхности
Поздрави на всички читатели на нашия блог Днес, скъпи читатели, искам да подчертая темата как да инсталирам гнезда. Тази процедура е много често търсена при подмяна на стар контакт с нов в случай на повреда, когато ...