Naukowcy policzyli liczbę neuronów w ludzkim mózgu. Elektryczne przejawy aktywności korowej
Kora półkul duży mózg reprezentuje najwyższą i najbardziej złożoną organizację Ośrodek nerwowy typ ekranu, którego działanie zapewnia regulację różnych funkcji organizmu i złożonych form zachowania.
Korę tworzy warstwa istoty szarej o grubości 3-5 mm. Istota szara zawiera komórki nerwowe (ponad 10 miliardów neuronów), włókna nerwowe i komórki neuroglejowe. Jego różne odcinki, różniące się od siebie pewnymi cechami lokalizacji i budowy komórek, układem włókien i znaczeniem funkcjonalnym, nazywane są polami, które po raz pierwszy opisał niemiecki lekarz i naukowiec K. Brodmann.
Cytoarchitektura
Wśród wielobiegunowych neuronów kory wyróżnia się komórki piramidalne, gwiaździste, wrzecionowate, pajęczakowe, poziome, „kandelabrowe”, komórki z podwójnym bukietem dendrytów i niektóre inne typy neuronów.
Neurony piramidalne stanowią podstawową i najbardziej specyficzną formę kory mózgowej. Mają wydłużony korpus w kształcie stożka, którego wierzchołek jest skierowany w stronę powierzchni kory. Dendryty rozciągają się od wierzchołków i bocznych powierzchni ciała. Aksony wychodzą z podstawy komórek piramidalnych.
Komórki piramidalne różnych warstw kory różnią się wielkością i mają inny wygląd wartość funkcjonalna. Małe komórki to interneurony. W tworzeniu biorą udział aksony dużych piramid silnikścieżki piramid.
Neurony kory znajdują się w niejasno odgraniczonych warstwach, które są oznaczone cyframi rzymskimi i ponumerowane od zewnątrz do wewnątrz. Każda warstwa charakteryzuje się przewagą jednego typu komórek. W korze mózgowej wyróżnia się sześć głównych warstw:
- I - molekularny;
- II - granulat zewnętrzny;
- III - piramidalny;
- IV - wewnętrzny granulat;
- V - zwojowy;
- VI - warstwa komórek polimorficznych.
I - Molekularny warstwa kory zawiera nie duża liczba małe asocjacyjne poziome komórki Cajala. Ich aksony biegną równolegle do powierzchni mózgu jako część splotu stycznego włókien nerwowych warstwy molekularnej. Jednak większość włókien tego splotu jest reprezentowana przez rozgałęzienia dendrytów leżących poniżej warstw.
II- Zewnętrzny granulat warstwę tworzą liczne małe neurony piramidalne i gwiaździste. Dendryty tych komórek wznoszą się do warstwy molekularnej, do której albo wchodzą aksony Biała materia lub tworząc łuki, wchodzą również do splotu stycznego włókien warstwy molekularnej.
III - Najszersza warstwa kory mózgowej - piramidalny. Zawiera neurony piramidalne, komórki Martinottiego i komórki wrzecionowe z podwójnym bukietem dendrytów. Szczytowe dendryty piramid sięgają do warstwy molekularnej, a boczne dendryty tworzą synapsy z sąsiednimi komórkami tej warstwy. Akson komórki piramidalnej zawsze rozciąga się od jej podstawy. W małych komórkach pozostaje w korze mózgowej, w dużych komórkach tworzy włókno mielinowe, które trafia do istoty białej mózgu. Aksony małych wielokątnych komórek Martinottiego są skierowane do warstwy molekularnej. Warstwa piramidalna pełni przede wszystkim funkcje asocjacyjne.
IV - Wewnątrz ziarnisty warstwa w niektórych obszarach kory jest bardzo rozwinięta (na przykład w obszarach wzrokowych i słuchowych kory), podczas gdy w innych może być prawie nieobecna (na przykład w zakręcie przedśrodkowym). Warstwa ta jest utworzona przez małe neurony gwiaździste. Zawiera dużą liczbę włókien poziomych.
V- Ganglionowy warstwę kory tworzą duże piramidy, a obszar kory ruchowej (zakręt przedśrodkowy) zawiera gigantyczne piramidy , które po raz pierwszy opisał kijowski anatom V. A. Bets. Szczytowe dendryty piramid sięgają pierwszej warstwy. Aksony piramid wystają do jąder motorycznych mózgu i rdzeń kręgowy. Najdłuższe aksony komórek Betza w drogach piramidowych docierają do ogonowych odcinków rdzenia kręgowego.
Oprócz neuronów piramidalnych warstwa zwojowa kory zawiera pionowe komórki wrzecionowate, których aksony wznoszą się do 1. warstwa kora mózgowa i komórki koszyczkowe.
VI - Warstwa komórki polimorficzne utworzone przez neurony o różnych kształtach (wrzecionowate, gwiaździste, komórki Martinottiego). Aksony tych komórek sięgają do istoty białej jako część dróg odprowadzających, a dendryty docierają do warstwy molekularnej.
Mieloarchitektura
Wśród włókien nerwowych kory mózgowej możemy wyróżnić asocjacyjny włókna łączące poszczególne obszary kory jednej półkuli, komisowy, łączący korę różnych półkul i występ włókna doprowadzające i odprowadzające, które łączą korę z jądrami w dolnych partiach ośrodkowego układu nerwowego. Włókna projekcyjne w korze mózgowej tworzą promienie promieniowe kończące się w trzeciej warstwie piramidalnej. Oprócz opisanego już splotu stycznego warstwy I - molekularnej, na poziomie IV - wewnętrznej warstwy ziarnistej i V - zwojowej znajdują się dwie warstwy styczne włókien nerwowych mielinowych - odpowiednio zewnętrzny pas Baillargera i wewnętrzny pas Baillargera. Dwa ostatnie układy to sploty utworzone przez końcowe odcinki włókien doprowadzających.
Modułowa zasada organizacji kory mózgowej
W korze mózgowej opisano powtarzające się bloki lub moduły neuronów, które uważa się za jej jednostki morfofunkcjonalne, zdolne do stosunkowo autonomicznej aktywności. Mają kształt cylindrów lub kolumn, biegnących pionowo przez całą grubość kory.
Każdy moduł obejmuje ścieżki doprowadzające, system połączeń lokalnych i ścieżki odprowadzające.
DO drogi aferentne obejmują włókna korowo-korowe i wzgórzowo-korowe.
Moduł jest zorganizowany wokół włókien korowo-korowych, które są aksonami komórek piramidalnych tej samej lub przeciwnej półkuli. Włókna korowo-korowe tworzą zakończenia we wszystkich warstwach kory danego modułu.
Moduł obejmuje również włókna wzgórzowo-korowe kończące się w warstwie IV kory na neuronach kolczastych gwiaździstych i dendrytach podstawnych neuronów piramidowych.
Ścieżki efektowe są utworzone przez aksony dużych i gigantycznych neuronów piramidalnych, a także aksony wrzecionowate i niektóre inne komórki warstwy VI kory.
System połączeń lokalnych jest tworzony przez interneurony modułu, które obejmują kilkanaście typów komórek. Większość z nich ma działanie hamujące i reguluje aktywność głównie neuronów piramidowych.
Z neuronów hamujących modułu najwyższa wartość Posiadać:
- komórki aksoaksonalne;
- ogniwa „kandelabrowe”;
- komórki koszykowe;
- komórki z podwójnym bukietem dendrytów;
- komórki za pomocą szczoteczki aksonalnej.
Układ neuronów hamujących pełni rolę filtra, hamując część neuronów piramidalnych kory.
Opony mózgowe i rdzeń kręgowy
Mózg i rdzeń kręgowy pokryte są trzema błonami tkanki łącznej: miękką, bezpośrednio przylegającą do tkanki mózgowej, pajęczynową i twardą, która graniczy tkanka kostna czaszka i kręgosłup. Opony zapewniają ochronę m.in. funkcja amortyzująca, zapewnia produkcję i wchłanianie płynu mózgowo-rdzeniowego.
Matko Pia bezpośrednio przylega do tkanki mózgowej i jest od niej ograniczony przez brzeżną błonę glejową. W luźnym włóknie tkanka łączna Powłoka zawiera dużą liczbę naczyń krwionośnych zaopatrujących mózg, liczne włókna nerwowe, aparat końcowy i pojedyncze komórki nerwowe. Pia mater otacza naczynia penetrujące mózg, tworząc wokół nich błonę okołonaczyniową. W komorach mózgu pia mater wraz z wyściółką bierze udział w tworzeniu splotów naczyniówkowych wytwarzających płyn mózgowo-rdzeniowy.
Pajęczynówka reprezentowana przez cienką warstwę luźnej włóknistej tkanki łącznej. Pomiędzy nim a pia mater znajduje się sieć poprzeczek składających się z cienkich wiązek kolagenu i cienkich elastycznych włókien. Sieć ta łączy ze sobą powłoki. Pomiędzy pia mater, która następuje po odciążeniu tkanki mózgowej, a pajęczynówką, która biegnie wzdłuż wzniesionych obszarów bez wchodzenia w zakamarki, znajduje się przestrzeń podpajęczynówkowa (podpajęczynówkowa), przesiąknięta cienkimi włóknami kolagenowymi i elastycznymi, które łączą membrany ze sobą Inny. Przestrzeń podpajęczynówkowa komunikuje się z komorami mózgu i zawiera płyn mózgowo-rdzeniowy. W tej przestrzeni są duże naczynia krwionośne, których gałęzie zasilają mózg.
Kosmki błony pajęczynówkowej (największe nazywane są ziarnistościami Pachionianu) służą jako miejsca, przez które substancje z płynu mózgowo-rdzeniowego wracają do krwi. Są to jałowe wyrostki błony pajęczynówkowej, zawierające sieć szczelinowatych przestrzeni i wystające do światła zatok opony twardej.
Dura mater utworzony przez gęstą włóknistą tkankę łączną zawierającą wiele elastycznych włókien. W jamie czaszki jest ściśle połączona z okostną. W kanał kręgowy opona twarda jest oddzielona od okostnej kręgu przestrzenią nadtwardówkową, wypełnioną warstwą luźnej włóknistej tkanki łącznej, która zapewnia jej pewną ruchliwość. Pomiędzy oponą twardą a błoną pajęczynówkową znajduje się przestrzeń podtwardówkowa. Przestrzeń podtwardówkowa zawiera niewielką ilość płynu.
Błony po stronie przestrzeni podtwardówkowej i podpajęczynówkowej pokryte są warstwą komórki płaskie charakter glejowy.
Zmiany związane z wiekiem
Zmiany w ośrodkowym układzie nerwowym podeszły wiek związane przede wszystkim ze zmianami sklerotycznymi w naczyniach krwionośnych mózgu. Na starość pia mater i błona pajęczynówkowa mózgu pogrubiają się. Mogą pojawiać się w nich osady wapienne. Obserwuje się zanik kory mózgowej półkule mózgowe głównie płat czołowy i ciemieniowy. Liczba neuronów na jednostkę objętości kory maleje, zależy to głównie od śmierci komórek. Neurony zmniejszają się, częściowo tracą substancję zasadochłonną, jądra stają się gęstsze, a ich zarys staje się nierówny. Piramidy warstwy V kory ruchowej i komórki gruszkowate kory móżdżku zmieniają się szybciej niż inne. Granulki lipofuscyny gromadzą się w neuronach różnych części układu nerwowego.
Wszystkie najważniejsze reakcje behawioralne człowieka zachodzą za pomocą centralnego układu nerwowego. Główne funkcje ośrodkowego układu nerwowego to:
- zjednoczenie wszystkich części ciała w jedną całość i ich regulacja;
- zarządzanie stanem i zachowaniem ciało zgodnie z warunkami otoczenie zewnętrzne i jego potrzeby.
U zwierząt wyższych i ludzi Wiodącą częścią ośrodkowego układu nerwowego jest kora mózgowa . Kontroluje najbardziej złożone funkcje w życiu człowieka procesy mentalne(świadomość, myślenie, mowa, pamięć itp.).
Głównymi metodami badania funkcji ośrodkowego układu nerwowego są metody usuwania i podrażnienia (w klinice i u zwierząt), rejestracja zjawisk elektrycznych, metoda odruchy warunkowe.
Wciąż rozwijane są nowe metody badania ośrodkowego układu nerwowego: wykorzystujące tzw tomografia komputerowa widać zmiany morfofunkcjonalne w mózgu na różnych głębokościach; fotografia w promieniach podczerwonych (obrazowanie termowizyjne) pozwala wykryć „najgorętsze” punkty w mózgu; Nowych danych na temat funkcjonowania mózgu dostarcza badanie jego oscylacji magnetycznych.
Podstawowe funkcje i oddziaływania neuronów
Głównymi elementami strukturalnymi układu nerwowego są komórki nerwowe lub neurony.
Podstawowe funkcje neuronów
Za pośrednictwem neuronów informacje przekazywane są z jednej części układu nerwowego do drugiej, następuje wymiana informacji pomiędzy układem nerwowym a różnymi częściami ciała. Najbardziej złożone procesy przetwarzania informacji zachodzą w neuronach. Z ich pomocą powstają odpowiedzi ciała (odruchy) na bodźce zewnętrzne i wewnętrzne.
Zatem, głównymi funkcjami neuronów są: odbieranie bodźców zewnętrznych funkcja receptora, ich przetwarzanie funkcję integracyjną i przekazywanie wpływów nerwowych na inne neurony lub różne pracujące narządy funkcja efektorowa. Główne procesy przetwarzania informacji zachodzą w ciele komórki nerwowej, zwanej somą. Liczne, rozgałęzione pędy przypominające drzewo dendryty(greckie drzewo dendronowy) służą jako wejścia neuronowe, przez które sygnały dostają się do komórki nerwowej. Wyjście neuronu jest procesem wychodzącym z ciała komórki akson(oś osi greckiej), która przekazuje impulsy nerwowe dalej do innej komórki nerwowej lub narządu roboczego (mięsień, gruczoł). Szczególnie silnie pobudliwa jest początkowa część aksonu i jego przedłużenie w miejscu wyjścia z ciała komórki. wzgórek aksonu neuronu. To właśnie w tym segmencie komórki powstaje impuls nerwowy.
Rodzaje neuronów
Neurony dzielą się na trzy główne typy: doprowadzające, odprowadzające i pośrednie. Neurony doprowadzające(wrażliwe lub dośrodkowe) przekazują informacje z receptorów do 1CNS. Ciała tych neuronów znajdują się poza OUN węzły kręgosłupa i w węzłach nerwy czaszkowe. Neurony doprowadzające mają długi proces - dendryt, który styka się na obwodzie z formacją percepcyjną - receptor lub sam tworzy receptor, a także drugi proces - akson, wchodzący przez tylne rogi w rdzeń kręgowy.
Neurony efektywne(odśrodkowe) są związane z przenoszeniem zstępujących wpływów z leżących poniżej pięter układu nerwowego na leżące poniżej lub z ośrodkowego układu nerwowego na narządy robocze. Neurony odprowadzające charakteryzują się rozgałęzioną siecią krótkich procesów - dendrytów i jednym długim procesem - aksonem.
Neurony pośrednie(interneurony lub interneurony) są to z reguły mniejsze komórki, które komunikują się między różnymi neuronami (w szczególności doprowadzającymi i odprowadzającymi). Przekazują wpływy nerwowe w kierunku poziomym (na przykład w obrębie jednego odcinka rdzenia kręgowego) i w kierunku pionowym (na przykład z jednego odcinka rdzenia kręgowego do innych segmentów położonych powyżej lub poniżej). Ze względu na liczne gałęzie aksonu interneurony mogą jednocześnie pobudzić dużą liczbę innych neuronów.
Synapsy pobudzające i hamujące
Następuje interakcja neuronów ze sobą (i z narządami efektorowymi). Specjalna edukacja synapsy (gr. kontakt). Tworzą je końcowe gałęzie neuronu na ciele lub procesy innego neuronu. Im więcej synaps na komórce nerwowej, tym bardziej odczuwa ona różne podrażnienia, a co za tym idzie, tym szersza jest sfera wpływu na jej działanie i możliwość uczestniczenia w różnych reakcjach organizmu. Szczególnie dużo synaps znajduje się w wyższych partiach układu nerwowego, a dokładnie w neuronach o najbardziej złożonych funkcjach.
W strukturze synapsy wyróżnić można trzy elementy (ryc. 1):
- błona presynaptyczna, utworzony przez pogrubienie błony końcowej gałęzi aksonu;
- szczelina synaptyczna między neuronami;
- błona postsynaptyczna pogrubienie sąsiedniej powierzchni następnego neuronu.
Ryż. 1. Schemat synapsy
Przed. błona presynaptyczna,
Szybko. błona postsynaptyczna,
C bąbelki synoptyczne,
luka synoptyczna,
M mitochondria,
Ach, acetylocholina
Receptory P i pory (Pory)
dendryt (D) następny
neuronu.
Strzałka jednostronne przewodzenie wzbudzenia.
W większości przypadków przeniesienie wpływu z jednego neuronu na drugi odbywa się chemicznie. W presynaptycznej części kontaktu znajdują się pęcherzyki synaptyczne , które zawierają specjalne substancje mediatorzy lub pośredników. Mogą to być acetylocholina (w niektórych komórkach rdzenia kręgowego, w węzłach wegetatywnych), noradrenalina (w zakończeniach włókien nerwowych współczulnych, w podwzgórzu), niektóre aminokwasy itp. Impulsy nerwowe docierające do zakończeń aksonów powodują opróżnienie pęcherzyków synaptycznych i uwolnienie przekaźnika do szczeliny synaptycznej.
W zależności od charakteru wpływu na kolejną komórkę nerwową rozróżnia się synapsy pobudzające i hamujące.
W synapsach pobudzających mediatory (na przykład acetylocholina) wiążą się z określonymi makrocząsteczkami błony synaptycznej i powodują jej depolaryzację. W tym przypadku małe i krótkotrwałe (około 1 ms) wahanie potencjału błony w kierunku depolaryzacja Lub pobudzający potencjał postsynaptyczny(EPSP). Aby neuron mógł się pobudzić, EPSP musi osiągnąć poziom progowy. W tym celu przesunięcie depolaryzacji potencjału błonowego musi wynosić co najmniej 10 mV. Działanie mediatora jest bardzo krótkotrwałe (1-2 ms), po czym następuje jego rozbicie na nieskuteczne komponenty (np. Acetylocholina jest rozkładana przez enzym cholinoesterazę na cholinę i kwas octowy ) muł i jest ponownie wchłaniany przez zakończenia presynaptyczne (na przykład noradrenalinę).
W synapsy hamujące zawarte mediatory hamujące (Na przykład, kwas gamma-aminomasłowy ). Ich działanie na błonę postsynaptyczną powoduje zwiększenie uwalniania jonów potasu z komórki i wzrost polaryzacji błony. W tym przypadku krótkotrwałe wahania potencjału błonowego w kierunku hiperpolaryzacja hamujący potencjał postsynaptyczny(TPSP). W rezultacie komórka nerwowa zostaje zahamowana. Trudniej jest ją pobudzić niż w stanie pierwotnym. Będzie to wymagało silniejszej stymulacji, aby osiągnąć krytyczny poziom depolaryzacji.
Pojawienie się odpowiedzi impulsowej neuronu
Na błonie ciała i dendrytach komórki nerwowej znajdują się synapsy pobudzające i hamujące . W pewnych momentach część z nich może być nieaktywna, natomiast część aktywnie oddziałuje na sąsiednie obszary membrany. Ogólna zmiana potencjału błonowego neuronu jest wynikiem: złożona interakcja(integracja) lokalnych EPSP i IPSP wszystkich licznych aktywowanych synaps. Przy jednoczesnym wpływie synaps pobudzających i hamujących następuje sumowanie algebraiczne (tj. wzajemne odejmowanie) ich efektów. W takim przypadku wzbudzenie neuronu nastąpi tylko wtedy, gdy będzie suma pobudzających potencjałów postsynaptycznych więcej niż kwota hamowanie . Ten nadmiar musi mieć określoną wartość progową (około 10 mV). Tylko w tym przypadku pojawia się potencjał czynnościowy komórki. Należy zauważyć, że ogólnie pobudliwość neuronu zależy od jego wielkości: im mniejsza komórka, tym większa jej pobudliwość .
Wraz z pojawieniem się potencjału czynnościowego proces przewodzenia impuls nerwowy wzdłuż aksonu i przekazując go do kolejnego neuronu lub narządu pracującego, tj. realizowana jest funkcja efektorowa neuronu. Impuls nerwowy jest głównym środkiem komunikacji między neuronami.
Zatem, przekazywanie informacji w układzie nerwowym odbywa się za pomocą dwóch mechanizmów: elektrycznego (EPSP; IPSP; potencjał czynnościowy) i chemiczne (mediatorzy).
Cechy aktywności ośrodków nerwowych
Właściwości ośrodków nerwowych są w dużej mierze związane z charakterystyką przewodzenia impulsów nerwowych przez synapsy łączące różne komórki nerwowe.
Cechy wzbudzenia przez ośrodki nerwowe
Ośrodek nerwowy nazywany zbiorem komórek nerwowych niezbędnych do pełnienia funkcji. Ośrodki te reagują odpowiednimi reakcjami odruchowymi na stymulację zewnętrzną otrzymywaną od związanych z nimi receptorów. Komórki ośrodków nerwowych reagują także na bezpośrednie podrażnienie przez przepływające przez nie substancje znajdujące się w krwi (wpływy humoralne). W kompletnym organizmie istnieje ścisła koordynacja - koordynację ich działań.
Przewodzenie fali wzbudzenia z jednego neuronu do drugiego przez synapsę zachodzi w większości komórek nerwowych na drodze chemicznej za pomocą mediatora, a mediator jest zawarty tylko w presynaptycznej części synapsy i jest nieobecny w błonie postsynaptycznej. Dlatego ważna cecha przeprowadzanie wzbudzenia poprzez kontakty synoptyczne przewodzenie jednostronne wpływy nerwowe, co jest możliwe tylko od błony presynaptycznej do błony postsynaptycznej i jest niemożliwe w kierunku przeciwnym. Pod tym względem przepływ impulsów nerwowych w łuku odruchowym ma określony kierunek od neuronów doprowadzających do neuronów interkalarnych, a następnie do odprowadzających neuronów ruchowych lub neuronów autonomicznych.
Bardzo ważne w działaniu układu nerwowego ma inną cechę przewodzenia wzbudzenia przez synapsy powolne przewodzenie. Czas spędzony na procesach zachodzących od momentu zbliżenia się impulsu nerwowego do błony presynaptycznej do momentu pojawienia się potencjałów w błonie postsynaptycznej nazywa się opóźnienie synaptyczne. W większości neuronów centralnych jest to około 0,3 ms. Następnie potrzeba więcej czasu na rozwój pobudzającego potencjału postsynaptycznego (EPSP) i potencjału czynnościowego. Cały proces przekazywania impulsu nerwowego (od potencjału czynnościowego jednej komórki do potencjału czynnościowego następnej komórki) przez jedną synapsę trwa około 1,5 ms. Wraz ze zmęczeniem, ochłodzeniem i wieloma innymi wpływami zwiększa się czas trwania opóźnienia synaptycznego. Jeśli jakakolwiek reakcja wymaga udziału duża liczba neuronów (wiele setek, a nawet tysięcy), wówczas całkowita wartość opóźnienia przewodzenia przez ośrodki nerwowe może wynosić dziesiąte części sekundy, a nawet całe sekundy.
Podczas aktywności odruchowej czas całkowity od momentu zastosowania podrażnienia zewnętrznego aż do pojawienia się reakcji organizmu tzw ukryty lub utajony czas odruchu zależy głównie od czasu przewodzenia przez synapsy. Ważna jest wielkość czasu utajonego odruchu wskaźnik stan funkcjonalny ośrodki nerwowe . Pomiar czasu utajonego prostej reakcji motorycznej człowieka na sygnał zewnętrzny jest szeroko stosowany w praktyce do oceny stanu funkcjonalnego ośrodkowego układu nerwowego (ryc. 2).
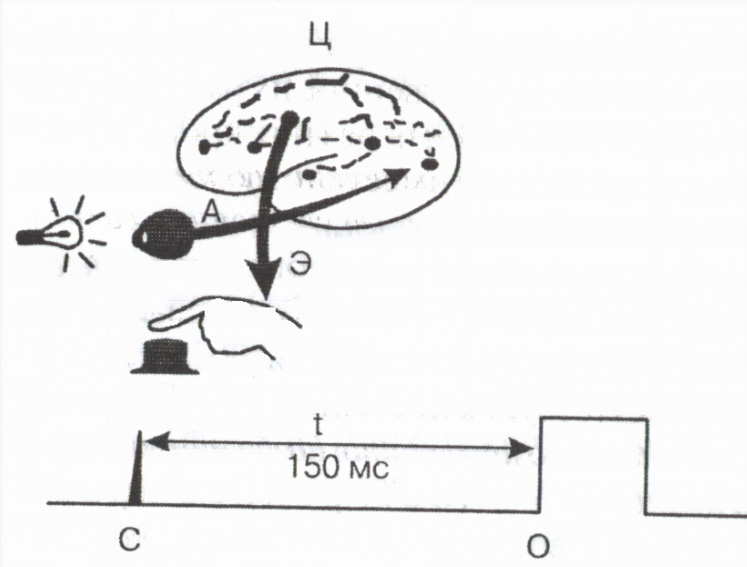
Ryż. 2. Schemat pomiaru czasu reakcji silnika
Doprowadzający
Drogi eferentne i C centralne; C - znak sygnału świetlnego,
Znak naciśnięcia przycisku O,
Czas reakcji 150ms.
Suma wzbudzeń
W odpowiedzi na pojedynczą falę doprowadzającą przemieszczającą się z receptorów do neuronów, w presynaptycznej części synapsy uwalniana jest niewielka ilość przekaźnika. W tym przypadku EPSP zwykle występuje w błonie postsynaptycznej neuronu, co stanowi niewielką lokalną depolaryzację. Aby całkowita wartość EPSP w całej błonie neuronu osiągnęła próg wystąpienia potencjału czynnościowego, wymagane jest zsumowanie wielu podprogowych EPSP na błonie komórkowej. Dopiero w wyniku takiego zsumowania wzbudzeń powstaje odpowiedź neuronu. Rozróżnia się sumowanie przestrzenne i czasowe.
Sumowanie przestrzenne obserwowane w przypadku jednoczesnego otrzymania kilku impulsów do tego samego neuronu wzdłuż różnych włókien presynaptycznych. Jednoczesne pobudzenie synaps w różnych częściach błony neuronowej zwiększa amplitudę całkowitego EPSP do wartości progowej. W rezultacie powstaje impuls odpowiedzi z neuronu i następuje reakcja odruchowa. Na przykład uzyskanie odpowiedzi z komórki motorycznej rdzenia kręgowego zazwyczaj wymaga jednoczesnej aktywacji 50–100 włókien doprowadzających z odpowiednich receptorów obwodowych.
Sumowanie czasu występuje, gdy ta sama droga aferentna jest aktywowana przez serię kolejnych bodźców. Jeżeli przerwy między przychodzącymi impulsami są wystarczająco krótkie, EPSP neuronu z poprzednich bodźców nie ma czasu na zanik, wówczas kolejne EPSP nakładają się na siebie, aż depolaryzacja błony neuronu osiągnie poziom krytyczny dla wystąpienia działania potencjał. W ten sposób nawet łagodne podrażnienia po pewnym czasie mogą powodować reakcje organizmu (np. kichanie i kaszel w odpowiedzi na łagodne podrażnienia błony śluzowej dróg oddechowych).
Transformacja i asymilacja rytmu
Charakter wyładowania odpowiedzi neuronu zależy nie tylko od właściwości bodźca, ale także od stanu funkcjonalnego samego neuronu(jego ładunek błonowy, pobudliwość, labilność). Komórki nerwowe mają właściwość zmiany częstotliwości przekazywanych impulsów, tj. właściwość transformacji rytmu.
Kiedy neuron jest bardzo pobudliwy (na przykład po zażyciu kofeiny), zwiększona częstotliwość impulsów (animacja rytmu) i przy niskiej pobudliwości (na przykład ze zmęczeniem). zwalnianie rytmu , ponieważ należy zsumować kilka przychodzących impulsów, aby ostatecznie osiągnąć próg wystąpienia potencjału czynnościowego. Te zmiany częstotliwości impulsów mogą wzmocnić lub osłabić reakcje organizmu na bodźce zewnętrzne.
Dzięki rytmicznej stymulacji aktywność neuronu może dostroić się do rytmu napływających impulsów, tj. zjawisko nabywania rytmu(Ukhtomsky A.A., 1928). Zapewnia rozwój nabywania rytmu dostrajanie aktywności wielu ośrodków nerwowych przy kontrolowaniu złożonych aktów motorycznych jest to szczególnie ważne dla utrzymania tempa ćwiczeń cyklicznych.
Śledzenie procesów
Po zakończeniu bodźca stan aktywny komórki nerwowej lub ośrodka nerwowego zwykle trwa przez pewien czas. Czas trwania procesów śladowych jest zróżnicowany: krótki w rdzeniu kręgowym (kilka sekund lub minut), znacznie dłuższy w ośrodkach mózgu (dziesiątki minut, godzin, a nawet dni) i bardzo długi w korze mózgowej (do kilkudziesięciu lat). .
Impulsy krążące w zamkniętych obwodach neuronów potrafią utrzymać wyraźny i krótkotrwały stan pobudzenia w ośrodku nerwowym. Długotrwałe ukryte ślady mają znacznie bardziej złożony charakter. Zakłada się, że długotrwałe zachowanie śladów w komórce nerwowej wszystkich charakterystyczne właściwości bodziec opiera się na zmianie struktury białek tworzących komórkę i restrukturyzacji kontaktów synaptycznych.
Następstwa krótkoimpulsowe (trwające do 1 godziny) są podstawą tzw pamięć krótkotrwała Podstawą powstawania są długoterminowe ślady związane z przegrupowaniami strukturalnymi i biochemicznymi w komórkach pamięć długoterminowa.
Koordynacja czynności ośrodkowego układu nerwowego
Procesy koordynacji czynności ośrodkowego układu nerwowego opierają się na koordynacji dwóch procesów nerwowych: pobudzenia i hamowania. Hamowanie jest aktywnym procesem neuronowym , co zapobiega lub tłumi podniecenie.
Znaczenie procesu hamowania w ośrodkowym układzie nerwowym
Zjawisko hamowania ośrodków nerwowych po raz pierwszy odkrył I.M. Sechenov w 1862 roku. Znaczenie tego procesu omówił on w książce „Reflexes of the Brain” (1863).
Zanurzając żabią udkę w kwasie i jednocześnie podrażniając niektóre części mózgu (np. nakładając kryształ sól kuchenna do obszaru międzymózgowia) I.M. Sechenov zaobserwował gwałtowne opóźnienie, a nawet całkowity brak odruchu „kwaśnego” rdzenia kręgowego (cofanie łapy). Na tej podstawie doszedł do wniosku, że niektóre ośrodki nerwowe mogą znacząco zmieniać aktywność odruchową innych ośrodków, w szczególności leżące nad nimi ośrodki nerwowe mogą hamować aktywność niższych. Opisane doświadczenie przeszło do historii fizjologii pod nazwą Hamowanie Sieczenowa .
Procesy hamujące są niezbędnym elementem koordynacji aktywności nerwowej. Po pierwsze, proces hamowania ogranicza rozprzestrzenianie się wzbudzenia na sąsiednie ośrodki nerwowe, co przyczynia się do jego koncentracji w niezbędnych obszarach układu nerwowego. Po drugie, powstające w niektórych ośrodkach nerwowych równolegle z pobudzeniem innych ośrodków nerwowych, następuje w ten sposób proces hamowania wyłącza niepotrzebne czynności ten moment narządy . Po trzecie, rozwój zahamowania w ośrodkach nerwowych chroni je przed nadmiernym przeciążeniem podczas pracy, tj. pełni rolę ochronną.
Hamowanie postsynaptyczne i presynaptyczne
Proces hamowania w przeciwieństwie do podniecenia, nie może rozprzestrzeniać się wzdłuż włókna nerwowego jest to zawsze proces lokalny w obszarze kontaktów synaptycznych. W zależności od miejsca pochodzenia wyróżnia się hamowanie presynaptyczne i postsynaptyczne.
Hamowanie postsynaptyczne są to efekty hamujące zachodzące w błonie postsynaptycznej. Najczęściej ten rodzaj hamowania jest związany z obecnością w ośrodkowym układzie nerwowym specjalnych neurony hamujące. Stanowią specjalny rodzaj interneuronów, w których zakończenia aksonów uwalniają przekaźnik hamujący. Jednym z takich mediatorów jest kwas gamma-aminomasłowy (GAM K.).
Impulsy nerwowe docierając do neuronów hamujących powodują w nich taki sam proces pobudzenia jak w innych komórkach nerwowych. W odpowiedzi normalny potencjał czynnościowy rozprzestrzenia się wzdłuż aksonu komórki hamującej. Jednakże, w przeciwieństwie do innych neuronów, zakończenia aksonów nie uwalniają przekaźnika pobudzającego, ale hamującego. W rezultacie komórki hamujące hamują te neurony, na których kończą się ich aksony.
Specjalne neurony hamujące obejmują komórki Renshawa w rdzeniu kręgowym, komórki Purkinjego w móżdżku, komórki koszyczkowe w międzymózgowiu itp. Przykładowo komórki hamujące odgrywają ogromne znaczenie w regulacji aktywności mięśni antagonistycznych: prowadząc do rozluźnienia mięśni antagonistycznych, ułatwiają tym samym równoczesny skurcz mięśni agonistycznych (ryc. 3).
Komórki Renshawa biorą udział w regulacji poziomu aktywności poszczególnych neuronów ruchowych rdzenia kręgowego. Kiedy neuron ruchowy jest wzbudzony, impulsy przemieszczają się wzdłuż jego aksonu do włókien mięśniowych i jednocześnie wzdłuż zabezpieczeń aksonu do komórki hamującej Renshawa. Aksony tego ostatniego „wracają” do tego samego neuronu, powodując jego zahamowanie. Im więcej impulsów pobudzających neuron ruchowy wysyła na obwód (a tym samym do komórki hamującej), tym silniejszy jest ten impuls. hamowanie powrotne(rodzaj hamowania postsynaptycznego). Taki zamknięty system działa jako mechanizm samoregulacji neuronu, chroniąc go przed nadmierną aktywnością.
Komórki Purkiniego móżdżku dzięki hamującemu działaniu na komórki jąder podkorowych i struktur macierzystych uczestniczą w regulacji napięcia mięśniowego.
Komórki koszykowe w międzymózgowiu są jak bramy, które pozwalają lub nie pozwalają impulsom docierającym do kory mózgowej z różnych obszarów ciała.
Hamowanie presynaptyczne występuje przed kontaktem synaptycznym w regionie presynadycznym. Zakończenie aksonu hamującej komórki nerwowej tworzy synapsę na końcu aksonu pobudzającej komórki nerwowej, powodując nadmiernie silną depolaryzację błony tego aksonu, co hamuje przepływające tu potencjały czynnościowe i tym samym blokuje przekazywanie pobudzenie. Ten rodzaj hamowania ogranicza przepływ impulsów doprowadzających do ośrodków nerwowych, wyłączając wpływy zewnętrzne w stosunku do głównej aktywności.
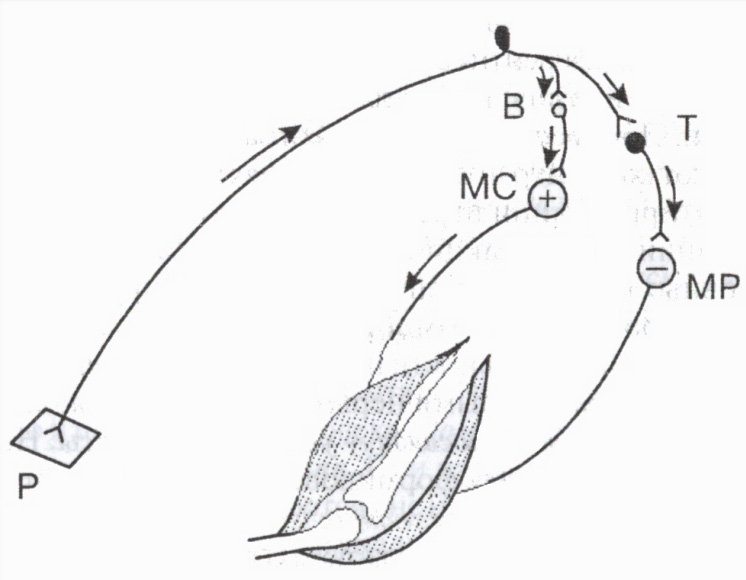
Ryż. 3. Udział komórki hamującej w regulacji pracy mięśni antagonistycznych
B i T to neurony pobudzające i hamujące. Pobudzenie (+) neuronu ruchowego mięśnia zginacza (MS) i hamowanie (-) neuronu ruchowego mięśnia prostownika (MR). Receptor skórny P.
Zjawisko napromieniowania i koncentracji
Gdy pobudzany jest jeden receptor, pobudzenie może w zasadzie rozprzestrzenić się w ośrodkowym układzie nerwowym w dowolnym kierunku i do dowolnej komórki nerwowej. Dzieje się tak z powodu licznych połączeń neuronów jednego łuk odruchowy z neuronami innych łuków odruchowych. Zjawisko to nazywa się rozprzestrzenianiem się procesu wzbudzenia na inne ośrodki nerwowe naświetlanie.
Im silniejsza jest stymulacja aferentna i im większa pobudliwość otaczających neuronów, tym więcej neuronów obejmuje proces napromieniania. Procesy hamowania ograniczają napromienianie i przyczyniają się do koncentracji wzbudzenia w punkcie początkowym ośrodkowego układu nerwowego.
Istotny jest proces napromieniania pozytywną rolę podczas powstawania nowych reakcji organizmu (reakcje orientacyjne, odruchy warunkowe). Im więcej różnych ośrodków nerwowych zostanie pobudzonych, tym łatwiej wybrać spośród nich te, które są najbardziej potrzebne do dalszych czynności. Dzięki naświetlaniu pobudzenie pomiędzy różnymi ośrodkami nerwowymi nowe relacje funkcjonalne powstają odruchy warunkowe . Na tej podstawie możliwe jest na przykład kształtowanie nowych umiejętności motorycznych.
Jednocześnie może mieć również napromienianie wzbudzenia negatywny wpływ na stan i zachowanie organizmu, zakłócając subtelne relacje pomiędzy pobudzonymi i hamowanymi ośrodkami nerwowymi oraz powodując zaburzona koordynacja ruchów .
Dominujący
Badając cechy relacji międzycentralnych, A. A. Ukhtomsky odkrył, że jeśli w ciele zwierzęcia zostanie przeprowadzona złożona reakcja odruchowa, na przykład powtarzające się akty połykania, wówczas elektryczna stymulacja ośrodków motorycznych nie tylko przestaje powodować ruch kończyn w tym chwili, ale także przyspiesza przebieg rozpoczętej reakcji łańcuchowej, czyli połykania, która okazała się dominująca.
Takie dominujące ognisko pobudzenia w ośrodkowym układzie nerwowym, które warunkuje bieżącą aktywność organizmu, A. A. Ukhtomsky (1.923) określił terminem dominującym.
Dominujące skupienie może wystąpić, gdy podwyższony poziom pobudliwość komórek nerwowych, która jest tworzona przez różne humoralne i wpływy nerwowe. Tłumi aktywność innych ośrodków, wywierając związane z tym hamowanie.
Zjednoczenie dużej liczby neuronów w jeden dominujący układ następuje poprzez wzajemne dostrojenie się do ogólnego tempa aktywności, czyli poprzez asymilację rytmu. Niektóre komórki nerwowe zmniejszają swoją wyższą aktywność, podczas gdy inne zwiększają swoją niską aktywność do pewnego średniego, optymalnego rytmu. Dominant może przez długi czas pozostawać w stanie ukrytym, śladowym (dominant potencjalny). Po wznowieniu poprzedniego stanu lub poprzedniej sytuacji zewnętrznej, może pojawić się ponownie dominacja (aktualizacja dominanty). Na przykład w stanie przedstartowym aktywowane są wszystkie ośrodki nerwowe, które były częścią układu pracy podczas poprzedniego treningu, i odpowiednio wzmacniane są funkcje związane z pracą. Mentalna egzekucja ćwiczenia fizyczne lub reprezentacja ruchów odtwarza także dominującą roboczą, która zapewnia efekt treningowy przedstawiania ruchów i jest podstawą tzw. Treningu ideomotorycznego. Z pełnym relaksem (na przykład z trening autogenny) sportowcy dążą do wyeliminowania dominatów roboczych, co przyspiesza procesy rekonwalescencji.
Jako czynnik zachowania, dominujący jest kojarzony z najwyższym aktywność nerwowa i psychologia człowieka. Dominantą jest podłoże fizjologiczne akt uwagi. W obecności dominującej wiele wpływów środowiska zewnętrznego pozostaje poza naszą uwagą, ale te, które szczególnie nas interesują, są intensywniej wychwytywane i analizowane. Zatem dominujący jest potężnym czynnikiem w wyborze najbardziej znaczących biologicznie i społecznie bodźców.
Funkcje rdzenia kręgowego i podkorowych części mózgu
W centralnym układzie nerwowym jest więcej starożytnych segmentowy i ewolucyjnie młodszy suprasegmentalny części układu nerwowego. Do odcinków segmentowych zalicza się rdzeń kręgowy, rdzeń przedłużony i śródmózgowie, których odcinki regulują funkcje poszczególnych części ciała leżących na tym samym poziomie. Sekcje suprasegmentalne - międzymózgowie, móżdżek i kora mózgowa nie mają bezpośrednich połączeń z narządami ciała, ale kontrolują ich aktywność poprzez leżące poniżej sekcje segmentowe.
Rdzeń kręgowy
Rdzeń kręgowy jest najniższą i najstarszą częścią centralnego układu nerwowego . Istota szara ludzkiego rdzenia kręgowego zawiera około 13,5 miliona komórek nerwowych. Z nich większość (97%) to komórki pośrednie (interneurony lub interneurony) , które zapewniają złożone procesy koordynacyjne w rdzeniu kręgowym. Wśród neuronów ruchowych rdzenia kręgowego są duże neurony ruchowe alfa i małe neurony motoryczne gamma. Najgrubsze i najszybciej przewodzące włókna nerwów ruchowych odchodzą od neuronów ruchowych alfa, powodując skurcze szkieletowy włókna mięśniowe. Cienkie włókna neuronów ruchowych gamma nie powodują skurczu mięśni. Zbliżają się do pro-prioreceptorów wrzeciona mięśniowe i regulują ich wrażliwość.
Odruchy rdzenia kręgowego Można podzielić na silnik przeprowadzane przez neurony ruchowe alfa rogów przednich i wegetatywny przeprowadzana przez komórki doprowadzające rogów bocznych.
Neurony ruchowe rdzenia kręgowego unerwiają wszystkie mięśnie szkieletowe (z wyjątkiem mięśni twarzy) . Rdzeń kręgowy wykonuje elementarne odruchy motoryczne zgięcie i wyprost, rytmiczne, kroczące, powstałe w wyniku podrażnienia skóry lub proprioceptorów mięśni i ścięgien, a także wysyła stałe impulsy do mięśni, wspierając napięcie mięśniowe. Specjalne neurony ruchowe unerwiają mięśnie oddechowe - mięśnie międzyżebrowe i przeponę i zapewniają ruchy oddechowe. Neurony autonomiczne unerwiają wszystkie narządy wewnętrzne (serce, naczynia krwionośne, gruczoły potowe, gruczoły wydzielina wewnętrzna, przewód pokarmowy, układ moczowo-płciowy).
Funkcja przewodnika Rdzeń kręgowy jest związany z przekazywaniem przepływu informacji otrzymywanych z obwodu do leżących nad nim części układu nerwowego oraz z przewodzeniem impulsów dochodzących z mózgu do rdzenia kręgowego.
Za ostatnie lata rozwinięty specjalne techniki do badania aktywności rdzenia kręgowego zdrowa osoba. Przykładowo stan funkcjonalny neuronów ruchowych alfa ocenia się na podstawie zmian potencjałów odpowiedzi mięśni podczas stymulacji obwodowej – tzw. Odruch H (odruch Hoffmanna) mięsień łydki kiedy jest zirytowany nerw piszczelowy i przez Odruch T (ze ścięgna ścięgna) mięśnia płaszczkowatego z podrażnieniem ścięgna Achillesa. Opracowano metody rejestrowania (z nienaruszonych powierzchni ciała) potencjałów przechodzących przez rdzeń kręgowy do mózgu.
Rdzeń przedłużony i most
Rdzeń przedłużony i most (razem tyłomózgowie) stanowią część pnia mózgu. Istnieje duża grupa nerwów czaszkowych (od par V do XII), które unerwiają skórę, błony śluzowe, mięśnie głowy i szereg narządy wewnętrzne(serce, płuca, wątroba). Tutaj są ośrodki wielu odruchów trawiennych żucie, połykanie, ruchy żołądka i części jelit, wydzielanie soków trawiennych, a także ośrodki niektórych odruchów obronnych (kichanie, kaszel, mruganie, łzawienie, wymioty) i ośrodki metabolizmu wody, soli i cukru . Na dnie komory IV, w rdzeniu przedłużonym, znajduje się ważny ośrodek oddechowy, składający się z ośrodków wdechu i wydechu. Składa się z małych komórek, które wysyłają impulsy do mięśni oddechowych za pośrednictwem neuronów ruchowych rdzenia kręgowego.
Położony w bliskiej odległości ośrodek sercowo-naczyniowy . Jego duże komórki regulują pracę serca i światło naczyń krwionośnych. Przeplatanie się komórek ośrodków oddechowych i sercowo-naczyniowych zapewnia ich ścisłą interakcję.
Rdzeń przedłużony odgrywa ważną rolę w realizacji aktów motorycznych oraz w regulacji napięcia mięśni szkieletowych, zwiększenie napięcia mięśni prostowników . Bierze udział w szczególności w realizacji odruchy regulacji postawy (szyjny, labirynt). Wznoszące się części wznoszące przechodzą przez rdzeń przedłużony ścieżki wrażliwości słuchowej, przedsionkowej, proprioceptywnej i dotykowej .
Śródmózgowie
Śródmózgowie składa się z jądra czworobocznego, istoty czarnej i jądra czerwonego. W przednich guzkach czworobocznych znajdują się wzrokowe ośrodki podkorowe i z tyłu słuchowy . Śródmózgowie jest zaangażowane regulacja ruchów oczu , wykonuje odruch źreniczny (rozszerzenie źrenic w ciemności i zwężenie w świetle).
Czworonogi wykonują szereg reakcji, którymi są elementy odruchu orientacyjnego . W reakcji na nagłe podrażnienie głowa i oczy zwracają się w stronę bodźca, a u zwierząt uszy sterczą. Odruch ten (według I.P. Pavlova odruch „Co to jest?”) jest niezbędny, aby przygotować organizm na szybką reakcję na każde nowe uderzenie.
Czarna substancja śródmózgowie jest związane z odruchami żucia i połykania regulacja napięcia mięśniowego (zwłaszcza gdy to robię małe ruchy palce) oraz w organizowaniu przyjaznych reakcji motorycznych.
Czerwony rdzeń śródmózgowie pełni funkcje motoryczne reguluje napięcie mięśni szkieletowych , powodując zwiększone napięcie mięśni zginaczy. Śródmózgowie, wywierając znaczący wpływ na napięcie mięśni szkieletowych, bierze udział w wielu odruchy instalacyjne służące utrzymaniu postawy (prostowanie ułożenia ciała koroną do góry itp.).
Międzymózgowie
Międzymózgowie obejmuje wzgórze (wzgórze wzrokowe) i podwzgórze (podwzgórze).
Poprzez wzgórze przechodzić przez wszystkie ścieżki doprowadzające (z wyjątkiem węchowych), które są wysyłane do odpowiednich obszarów percepcyjnych kory (słuchowej, wzrokowej itp.). Jądra wzgórza są podzielone na specyficzne i niespecyficzne . Konkretne obejmują Rdzenie przełączające (przekaźnikowe) i asocjacyjne . Wpływy doprowadzające ze wszystkich receptorów ciała są przekazywane przez jądra przełączające wzgórza. Jądra asocjacyjne otrzymują impulsy od jąder przełączających i zapewniają ich interakcję. Oprócz tych jąder wzgórze zawiera niespecyficzne jądra, które mają zarówno działanie aktywujące, jak i hamujące na małe obszary kory.
Dzięki rozbudowanym połączeniom wzgórze odgrywa istotną rolę w funkcjonowaniu organizmu. Impulsy dochodzące ze wzgórza do kory zmieniają stan neuronów korowych i regulują rytm aktywności korowej . Przy bezpośrednim udziale wzgórza następuje tworzenie odruchów warunkowych i rozwój umiejętności motorycznych, kształtowanie emocji osoba, jej wyraz twarzy. W szczególności wzgórze odgrywa dużą rolę w występowaniu wrażeń odczucia bólu . Jego działalność jest powiązana regulacja biorytmów w życiu człowieka (codziennie, sezonowo itp.).
Podwzgórze jest najwyższym podkorowym ośrodkiem regulacji funkcje wegetatywne , stwierdza czuwanie i sen . Znajdują się tu ośrodki wegetatywne, regulujące metabolizm i ciało, zapewniając utrzymywanie stałej temperatury ciała (u zwierząt stałocieplnych) i normalny poziom ciśnienie krwi , wspierający bilans wodny regulujący uczucie głodu i sytości . Podrażnienie tylnych jąder podwzgórza powoduje wzrost wpływów współczulnych, a przednich - przywspółczulnych.
Ze względu na połączenie podwzgórza z przysadką mózgową ( układ podwzgórzowo-przysadkowy) kontrolowana jest aktywność gruczołów dokrewnych. Reakcje autonomiczne i hormonalne, regulowane przez podwzgórze, są składową reakcji emocjonalnych i motorycznych człowieka.
Niespecyficzny układ mózgowy
System niespecyficzny zajmuje środkową część pnia mózgu. Nie polega na analizie jakiejś szczególnej wrażliwości i realizacji określonych reakcji odruchowych. Impulsy do tego układu dochodzą bocznymi gałęziami ze wszystkich określonych ścieżek, powodując ich rozległą interakcję. Dla system niespecyficzny Charakterystyczne jest umiejscowienie neuronów w postaci sieci rozproszonej, obfitość i różnorodność ich procesów. W związku z tym otrzymał nazwę formacja siatkowata lub formacja siatkowata.
Wyróżnić dwa rodzaje wpływów niespecyficzny układ na pracę innych ośrodków nerwowych aktywizujące i hamujące . Obydwa typy tych wpływów mogą mieć charakter rosnący (do ośrodków leżących niżej) i malejący (do ośrodków położonych niżej). Służą do regulacja stanu funkcjonalnego mózgu, poziomu czuwania oraz regulacja reakcji posturalno-tonicznych i fazowych mięśni szkieletowych.
Móżdżek
Móżdżek jest formacją suprasegmentalną, która nie ma bezpośredniego połączenia z aparatem wykonawczym. Móżdżek składa się z niesparowanej formacji - robaka i sparowanych półkul.
Główne neurony kory móżdżku są liczne Komórki Purkiniego. Dzięki rozbudowanym połączeniom (każda komórka ma aż 200 000 synaps) zawierają integracja szerokiej gamy wpływów sensorycznych , głównie proprioceptywne, dotykowe i przedsionkowe. Reprezentacja różnych receptorów obwodowych w korze móżdżku ma organizacja somatotopowa (gr. somatos body, topos miejsce), czyli odzwierciedla kolejność ich umiejscowienia w organizmie człowieka. Ponadto ten porządek ułożenia odpowiada temu samemu porządkowi ułożenia reprezentacji części ciała w korze mózgowej, co ułatwia wymianę informacji między korą a móżdżkiem i zapewnia ich wspólne działanie w sterowaniu ludzkim zachowaniem. Decyduje o tym prawidłowa organizacja geometryczna neuronów móżdżku znaczenie w liczeniu czasu i wyraźnym utrzymywaniu tempa ruchów cyklicznych.
Główną funkcją móżdżku jest regulacja reakcji posturalnych i koordynacja aktywności ruchowej.(Orbeli Los Angeles, 1926).
Przez cechy anatomiczne(połączenia kory móżdżku z jądrami) i znaczenie funkcjonalne, móżdżek dzieli się na trzy strefy podłużne:
- wewnętrzna lub środkowa kora robaka, którego zadaniem jest regulacja napięcia mięśni szkieletowych, utrzymanie postawy ciała i równowagi;
- mediator Środkowa cześć Kora mózgowa móżdżek, którego funkcją jest koordynacja reakcji posturalnych z ruchami i korygowanie błędów;
- kora boczna lub boczna półkul móżdżek, który wraz z międzymózgowie a kora mózgowa bierze udział w programowaniu szybkich ruchów balistycznych (rzuty, uderzenia, skoki itp.).
Zwoje podstawne
Do zwojów podstawy zalicza się prążkowie, składające się z jądra ogoniastego i skorupy oraz jądra bladego, i obecnie uważa się je również za migdał(związany z ośrodki wegetatywne układ limbiczny) i istotę czarną śródmózgowia.
Wpływy doprowadzające docierają do zwojów podstawy mózgu z receptorów ciała poprzez wzgórze i ze wszystkich obszarów kory mózgowej. Prawie wyłącznie dostają się do prążkowia. Wpływające z niego wpływy kierowane są do bladego jądra i dalej do ośrodków pnia układu pozapiramidowego, a także przez wzgórze z powrotem do kory.
Zwoje podstawy biorą udział w tworzeniu odruchów warunkowych i realizacji kompleksów odruchy bezwarunkowe(obronne, spożywcze itp.). Zapewniają niezbędną pozycję ciała podczas pracy fizycznej, a także przepływ automatycznych, rytmicznych ruchów (starożytne automatyzmy).
Jądro blade pełni główną funkcję motoryczną, a prążkowie reguluje jego aktywność. Obecnie odkryto znaczenie jądra ogoniastego w kontroli złożonych procesów umysłowych – uwagi, pamięci i wykrywania błędów.
Wszystkie funkcje organizmu można podzielić na somatyczne, Lub zwierzę(zwierzęta) związane z percepcją informacje zewnętrzne i aktywność mięśni oraz wegetatywna (roślina) związana z czynnością narządów wewnętrznych, procesami oddychania, krążenia krwi, trawieniem, wydalaniem, metabolizmem, wzrostem i rozmnażaniem.
Organizacja funkcjonalna autonomicznego układu nerwowego
Autonomiczny układ nerwowy zwany zespołem odprowadzających komórek nerwowych rdzenia kręgowego i mózgu, a także komórkami specjalnych węzłów (zwojów) unerwiających narządy wewnętrzne. Stymulacja różnych receptorów ciała może powodować zmiany zarówno w funkcjach somatycznych, jak i autonomicznych, ponieważ wspólne są części doprowadzające i środkowe tych łuków odruchowych. Różnią się jedynie odcinkami odprowadzającymi. Cecha charakterystyczna eferentnymi drogami wchodzącymi w skład łuków odruchowych odruchów autonomicznych jest ich dwuneuronowa struktura(jeden neuron znajduje się w ośrodkowym układzie nerwowym, drugi w zwojach lub w narządzie unerwionym).
Wegetatywny system nerwowy dzieli się na dwie części: współczulną i przywspółczulną (ryc. 4).
Ścieżki efektowe współczulny układ nerwowy zacznij od klatki piersiowej i rejony lędźwiowe rdzeń kręgowy z neuronów jego rogów bocznych. Przeniesienie wzbudzenia z przednodalnych włókien współczulnych na postnodalne następuje przy udziale mediatora acetylocholina oraz od włókien postnodalnych do narządów unerwionych przy udziale mediatora noradrenalina . Wyjątkiem są włókna unerwiające gruczoły potowe i rozszerzające naczynia mięśni szkieletowych, gdzie pobudzenie przekazywane jest za pomocą acetylocholiny.
Ścieżki efektowe przywspółczulny układ nerwowy zaczynają się w mózgu od niektórych jąder środkowych i rdzeń przedłużony oraz w rdzeniu kręgowym z neuronów rejon sakralny. Przewodzenie wzbudzenia w synapsach szlaku przywspółczulnego zachodzi przy udziale mediatora acetylocholina . Drugi neuron odprowadzający znajduje się w unerwionym narządzie lub w jego pobliżu.
Najwyższym regulatorem funkcji autonomicznych jest podwzgórze , który działa w połączeniu z tworzeniem siatkowym i układem limbicznym pod kontrolą kory mózgowej. Ponadto neurony zlokalizowane w samych narządach lub w węzły współczulne, mogą przeprowadzać własne reakcje odruchowe bez udziału ośrodkowego układu nerwowego „odruchy peryferyjne” .
Ryż. 4. Autonomiczny układ nerwowy
Obszar wyjściowy lewego włókna: przywspółczulny (czarny)
i układy współczulne (zacienione).
Po prawej stronie znajduje się struktura eferentnej części łuku odruchowego odruchów autonomicznych. Po lewej stronie znajduje się schemat środka, rdzenia przedłużonego i rdzenia kręgowego.
Cyfry arabskie to numery odcinków piersiowych, cyfry rzymskie to numery odcinków lędźwiowych.
Funkcje współczulnego układu nerwowego
Przy udziale współczulnego układu nerwowego w organizmie zachodzi wiele ważnych odruchów, mających na celu zapewnienie jego stanu aktywnego, w tym aktywności motorycznej. Należą do nich refleks rozszerzenie oskrzeli, przyspieszenie akcji serca i nasilenie , rozszerzenie naczyń krwionośnych serca i płuc przy jednoczesnym zwężeniu naczyń krwionośnych skóry i narządów Jama brzuszna (zapewnienie redystrybucji krwi ), uwolnienie zmagazynowanej krwi z wątroby i śledziony, rozkład glikogenu do glukozy w wątrobie ( mobilizacja węglowodanów, źródeł energii ), zwiększona aktywność gruczołów dokrewnych i potowych. Współczulny układ nerwowy zmniejsza aktywność wielu narządów wewnętrznych: w wyniku zwężenia naczyń w nerkach zmniejszają się procesy powstawania moczu, hamowana jest aktywność wydzielnicza i motoryczna narządów przewód pokarmowy; czynność oddawania moczu zostaje uniemożliwiona, mięsień ściany rozluźnia się Pęcherz moczowy i jego zwieracz się kurczy.
Zwiększonej aktywności organizmu towarzyszy układ współczulny odruch rozszerzenia źrenic . Ogromne znaczenie dla aktywności motorycznej organizmu ma Troficzny wpływ nerwów współczulnych na mięśnie szkieletowe poprawiając ich metabolizm i stan funkcjonalny, łagodząc zmęczenie.
Współczulny układ nerwowy nie tylko podnosi poziom funkcjonowania organizmu, ale także mobilizuje swoje ukryte rezerwy funkcjonalne , aktywuje aktywność mózgu, zwiększa reakcje obronne ( reakcje immunologiczne, mechanizmy barierowe itp.), wywołuje reakcje hormonalne. Szczególne znaczenie ma współczulny układ nerwowy podczas rozwoju warunki stresowe , w większości trudne warunki aktywność życiowa. – podkreślił L. A. Orbeli istotne znaczenie współczulne wpływy na adaptację (przystosowanie) ciała do ciężkiej pracy, do różne warunki otoczenie zewnętrzne. Funkcja ta została przez niego nazwana adaptacyjno-troficzny.
Funkcje przywspółczulnego układu nerwowego
Przywspółczulny układ nerwowy wykonuje zwężenie oskrzeli, spowolnienie i osłabienie skurczów serca: zwężenie naczyń krwionośnych serca; uzupełnienie zasobów energetycznych (synteza glikogenu w wątrobie i wzmocnienie procesów trawiennych); wzmocnienie procesów tworzenia moczu w nerkach i zapewnienie oddawania moczu (skurcz mięśni pęcherza i rozluźnienie zwieracza) itp. Przywspółczulny układ nerwowy zapewnia głównie wyzwalające wpływy : zwężenie źrenicy, oskrzeli, aktywacja aktywności gruczoły trawienne i tak dalej.
Aktywność przywspółczulnego podziału autonomicznego układu nerwowego jest skierowana na prąd regulacja stanu funkcjonalnego, aby zachować stałość środowisko wewnętrzne homeostaza . Zapewnia dział przywspółczulny przywrócenie różnych wskaźników fizjologicznych , gwałtownie się zmienił po intensywnym praca mięśni, uzupełnienie zużytych zasobów energii. Mediator układ przywspółczulny acetylocholina, zmniejszając wrażliwość receptorów adrenergicznych na działanie adrenaliny i noradrenaliny, ma pewne działanie efekt antystresowy .
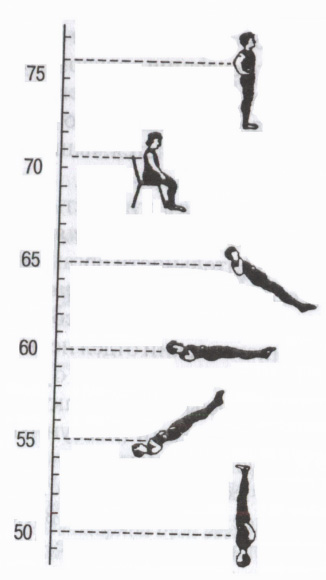
Ryż. 5. Odruchy autonomiczne
Wpływ pozycji ciała na tętno (bpm). (wg: Mogendovich M.R., 1972)
Poprzez autonomiczne szlaki współczulne i przywspółczulne centralny układ nerwowy realizuje pewne odruchy autonomiczne, zaczynając od różnych receptorów środowiska zewnętrznego i wewnętrznego: trzewno-trzewnego (od narządów wewnętrznych do narządów wewnętrznych - na przykład odruch oddechowo-sercowy); skórno-trzewny (ze skóry - zmiany czynności narządów wewnętrznych podczas podrażnienia aktywne punkty skóra, na przykład akupunktura, akupresura); z receptorów gałki ocznej odruchu oczno-sercowego Aschnera (zmniejszenie bicia serca przy naciskaniu gałki oczne efekt przywspółczulny); motoryczno-trzewny – np. próba ortostatyczna(zwiększone tętno podczas przechodzenia z pozycji leżącej do pozycji stojącej, efekt współczulny) itp. (ryc. 5). Służą do oceny stanu funkcjonalnego organizmu, a zwłaszcza stanu autonomicznego układu nerwowego (ocena wpływu jego układu współczulnego lub przywspółczulnego).
Układ limbiczny
Pod układ limbiczny rozumieć szereg struktur korowych i podkorowych, których funkcje są związane z organizacją reakcje motywacyjno-emocjonalne, pamięć i procesy uczenia się .
Sekcje korowe układu limbicznego, reprezentujące jego najwyższą sekcję, znajdują się na dolnych i wewnętrznych powierzchniach półkul mózgowych (części kory czołowej, zakrętu obręczy lub kory limbicznej, hipokampu itp.). DO struktury podkorowe Układ limbiczny obejmuje podwzgórze, niektóre jądra wzgórza, śródmózgowie i formację siatkową. Pomiędzy wszystkimi tymi działami istnieją ścisłe połączenia bezpośrednie i sprzężenia zwrotnego, tworząc tzw. „pierścień limbiczny”.
Układ limbiczny bierze udział w różnorodnych przejawach aktywności organizmu: w regulacji zachowań związanych z jedzeniem i piciem, cyklu snu i czuwania, w procesach tworzenia śladów pamięciowych (odkładanie i wydobywanie z pamięci), w rozwoju reakcji agresywno-obronnych , zapewniając selektywne zachowanie. Ona tworzy pozytywne i negatywne emocje ze wszystkimi komponentami motorycznymi, autonomicznymi i hormonalnymi. Elektryczna stymulacja różnych części układu limbicznego poprzez wszczepione elektrody (w doświadczeniach na zwierzętach, w klinice podczas leczenia pacjentów) ujawniła obecność centra przyjemności, wzbudzanie pozytywnych emocji i niezadowolenie, wywołując negatywne emocje. Pojedyncze podrażnienie takich punktów w głębokich strukturach ludzkiego mózgu powodowało pojawienie się uczuć „bezprzyczynowej radości”, „bezsensownej melancholii” i „niewytłumaczalnego strachu”.
Funkcje kory mózgowej
U wyższych ssaków i ludzi wiodącą częścią centralnego układu nerwowego jest kora mózgowa.
Neurony korowe
Kora jest warstwą istoty szarej o grubości 2-3 mm, zawierającą średnio około 14 miliardów komórek nerwowych. Cechą charakterystyczną jest obfitość połączeń międzyneuronowych, których rozwój trwa do 18. roku życia, a w niektórych przypadkach nawet dłużej.
Głównymi typami komórek korowych są neurony piramidalne i gwiaździste. Neurony gwiaździste związany z procesami percepcji podrażnień i unifikacją aktywności różnych neuronów piramidowych.
Neurony piramidalne pełnią funkcję eferentną kory (głównie poprzez przewód piramidowy) i procesy interakcji korowo-wiutrycznej pomiędzy neuronami oddalonymi od siebie. Największe komórki piramidalne, gigantyczne piramidy Betza, znajdują się w przednim centralnym zakręcie (obszar motoryczny kory).
Jednostką funkcjonalną kory jest kolumna pionowa wzajemnie połączone neurony. Pionowo wydłużone duże komórki piramidalne z neuronami znajdującymi się nad i pod nimi tworzą funkcjonalne stowarzyszenia neuronów. Masa Neurony kolumny pionowej odpowiadają na tę samą stymulację aferentną (z tego samego receptora) tą samą reakcją i wspólnie tworzą odpowiedzi eferentne neuronów piramidalnych. W razie potrzeby kolumny pionowe można łączyć w większe formacje, co pozwala na prowadzenie złożonych reakcji.
Znaczenie funkcjonalne różnych pól korowych
Według cech strukturalnych i znaczenia funkcjonalnego poszczególnych obszarów korowych cała skorupa jest podzielona na trzy główne grupy pól pierwotnych, wtórnych i trzeciorzędowych (ryc. 6).
Pola podstawowe związane z narządami zmysłów i narządami ruchu na obwodzie. Zapewniają pojawienie się wrażeń . Należą do nich na przykład pole bólu i wrażliwość mięśniowo-stawowa w tylnym środkowym zakręcie kory, pole widzenia w okolicy potylicznej, pole słuchowe w okolicy skroniowej i pole motoryczne w przednim zakręcie centralnym. W polach pierwotnych występują wysoce wyspecjalizowane komórki determinacyjne lub detektory, selektywnie reagując tylko na określone bodźce. Na przykład w korze wzrokowej znajdują się neurony detektorowe, które są wzbudzane tylko wtedy, gdy światło jest włączone lub wyłączone, wrażliwe tylko na określone natężenie, na określone interwały ekspozycji na światło, na określoną długość fali itp. Kiedy pierwotne pola kory ulegają zniszczeniu, dochodzi do tzw. ślepoty korowej, głuchoty korowej itp.
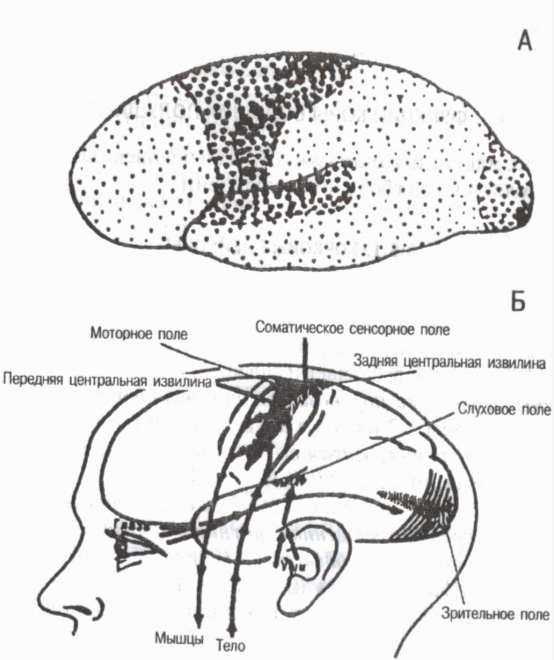
Ryż. 6. Pola pierwotne, wtórne i trzeciorzędne kory mózgowej
Na A: duże punkty pól pierwotnych, średnie pola wtórne, małe pola trzeciorzędowe (wg G. I. Polyakova 1964, A. R. Luria, 1971)
Na B: pierwotne (projekcyjne) pola kory mózgowej (wg V. Penfielda, L. Roberta, 1964)
Pola wtórne znajdują się obok głównych. Co się w nich dzieje rozumienia i rozpoznawania dźwięku, światła i innych sygnałów, powstają złożone formy uogólnionej percepcji . Kiedy pola wtórne ulegają uszkodzeniu, zdolność widzenia przedmiotów i słyszenia dźwięków zostaje zachowana, ale osoba ich nie rozpoznaje i nie pamięta ich znaczenia.
Pola trzeciorzędne rozwinęła się niemal wyłącznie u ludzi. Są to obszary asocjacyjne kory, zapewniające wyższe formy analizy i syntezy oraz kształtujące celową ludzką aktywność behawioralną. Pola trzeciorzędowe znajdują się w tylnej połowie kory w obszarach międzyciemieniowych, potylicznych i skroniowych oraz w przedniej połowie w częściach przednich obszary czołowe. Ich rola jest szczególnie duża w organizowaniu skoordynowanej pracy obu półkul . Pola trzeciorzędowe dojrzewają u ludzi później niż inne pola korowe i ulegają degradacji wcześniej niż inne podczas starzenia.
Funkcją tylnych pól trzeciorzędowych (głównie dolnych obszarów ciemieniowych kory) jest odbieranie, przetwarzanie i przechowywanie informacji. Tworzą wyobrażenie diagram ciała i diagram przestrzeni , zapewniając przestrzenną orientację ruchów. Przednie pola trzeciorzędowe (obszary czołowe) pełnią ogólną regulację złożonych form ludzkiego zachowania, formułowanie zamierzeń i planów, programów ruchów wolontariackich oraz monitorowanie ich realizacji . Związany jest z tym rozwój pól trzeciorzędowych u człowieka z funkcją mowy. Myślenie ( mowa wewnętrzna) jest możliwe tylko przy wspólnym działaniu różnych systemów sensorycznych, z których integracja informacji następuje w polach trzeciorzędnych. Przy wrodzonym niedorozwoju pól trzeciorzędowych osoba nie jest w stanie opanować mowy (wymawia tylko bezsensowne dźwięki), a nawet najprostszych umiejętności motorycznych (nie może się ubierać, używać narzędzi itp.).
Aktywność par i dominacja półkuli
W rezultacie następuje przetwarzanie informacji sparowana aktywność obu półkul mózg. Jednakże, co do zasady, jedna z półkul jest dominującą dominującą . Większość ludzi z liderem prawa ręka(praworęczny) Dominantą jest lewa półkula, a podrzędną (subdominantą) jest prawa półkula.
Lewa półkula w porównaniu z prawicą ma lepszą strukturę neuronową, większe bogactwo połączeń neuronowych, bardziej skoncentrowaną reprezentację funkcji i Lepsze warunki dopływ krwi W lewej półkuli dominującej znajduje się ośrodek mowy motorycznej (centrum Broki), który zapewnia aktywność mowy oraz zmysłowy ośrodek mowy, który rozumie słowa. Lewa półkula specjalizuje się w precyzyjnej kontroli sensomotorycznej ruchów dłoni.
U ludzi występują trzy formy asymetria funkcjonalna: motoryczne, sensoryczne i psychiczne . Zazwyczaj dana osoba ma dominującą rękę, nogę, oko i ucho. Problem asymetrii funkcjonalnej jest jednak dość złożony. Na przykład osoba praworęczna może mieć dominujące lewe oko lub lewe ucho, z których dominują sygnały. Co więcej, na każdej półkuli można reprezentować funkcje nie tylko przeciwnej, ale także tej samej strony ciała. Dzięki temu możliwe jest zastąpienie jednej półkuli inną w przypadku uszkodzenia, a także utworzenie strukturalne podstawy zmiennej dominacji półkuli w kontroli motorycznej.
Asymetria mentalna objawia się w postaci pewnej specjalizacje półkulowe. Dla lewa półkula charakteryzuje się procesami analitycznymi, sekwencyjnym przetwarzaniem informacji, w tym za pomocą mowy, myśleniem abstrakcyjnym, oceną tymczasowych relacji, przewidywaniem przyszłych wydarzeń, pomyślnym rozwiązywaniem problemów werbalnych i logicznych. W prawa półkula informacje przetwarzane są całościowo, syntetycznie (bez rozbijania na szczegóły), z uwzględnieniem przeszłych doświadczeń i bez udziału mowy, dominuje myślenie merytoryczne. Cechy te pozwalają powiązać percepcję cech przestrzennych i rozwiązywanie problemów wzrokowo-przestrzennych z prawą półkulą. Funkcje prawej półkuli są związane z czasem przeszłym, a lewej półkuli z przyszłością.
Aktywność elektryczna kory mózgowej
Zmiany stanu funkcjonalnego kory znajdują odzwierciedlenie w jej zapisie aktywność elektryczna elektroencefalogram (EEG). Nowoczesne elektroencefalografy wzmacniają potencjały mózgu 2-3 miliony razy i umożliwiają badanie EEG z wielu punktów kory jednocześnie, czyli badanie procesy systemowe. Rejestracja EEG produkowane w formie pisma tuszem na papierze, a także w formie pełny obraz na diagramie powierzchni mózgu, czyli mapach mózgu (metoda mapowania) na ekranie monitora współczesnych systemów komputerowych (ryc. 7).
Są pewne zakresy częstotliwości , zwany Rytmy EEG (ryc. 8): w stanie względnego spoczynku najczęściej rejestruje się rytm alfa (8-13 oscylacji na 1 s); w stanie aktywnej uwagi - rytm beta (14 oscylacji przy 1 s i powyżej); podczas zasypiania, niektórzy Stany emocjonalne rytm beta (4-7 oscylacji na 1 s); w głębokim śnie, utrata przytomności, znieczulenie, rytm delta (1-3 fluktuacje na 1 s).
EEG odzwierciedla cechy interakcji neuronów korowych podczas procesów psychicznych i Praca fizyczna (Livanov M.N., 1972). Brak ugruntowanej koordynacji podczas wykonywania nietypowej lub trudnej pracy prowadzi do tzw Desynchronizacja EEG szybka aktywność asynchroniczna. W miarę rozwoju umiejętności motorycznych w EEG pojawiają się zjawiska Synchronizacja EEG zwiększenie wzajemnych powiązań (synchronizacji i fazy) aktywności elektrycznej różnych obszarów kory zaangażowanych w kontrolowanie ruchów. Podczas pracy cyklicznej potencjały powolne pojawiają się w tempie wykonywanego, wyimaginowanego lub nadchodzącego ruchu „wyznaczone rytmy”(Sologub E.B., 1973).
Oprócz aktywności w tle w EEG, oddziel potencjały związane z dowolnymi zdarzeniami: potencjały wywołane, powstałe w odpowiedzi na bodźce zewnętrzne (słuchowe, wzrokowe itp.); potencjałów odzwierciedlających procesy mózgowe podczas przygotowania, realizacji i zakończenia poszczególnych aktów motorycznych „fala oczekiwań” lub uwarunkowana fala ujemna (Walter G., 1966), potencjały przedmotorowe, motoryczne i końcowe itp. Ponadto różne ultrawolne oscylacje trwające od kilku sekund do kilkudziesięciu minut (w szczególności tzw. „potencjały omega” itp.), które odzwierciedlają biochemiczne procesy regulacji funkcji i aktywności umysłowej.

Ryż. 7. Mapowanie mózgu
Wielokanałowy zapis ludzkiego elektroencefalogramu (EEG) na ekranie monitora i odbicie wzbudzonych (strefy jasne) i zahamowanych (strefy ciemne) obszarów kory mózgowej.
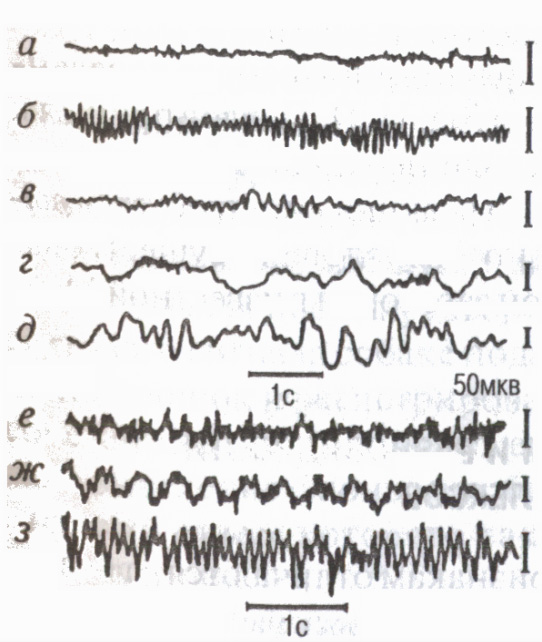
Ryż. 8. EEG obszarów potylicznych (a-e) i motorycznych (f-h) kory mózgowej człowieka podczas różne stany oraz podczas pracy mięśni
stan aktywny,
oczy otwarte (rytm beta); b- odpoczynek, oczy zamknięte (rytm alfa);
c senność (rytm tesha);
d zasypianie; D głęboki sen"H
(rytm delta); e niezwykłe lub
ciężka praca asynchroniczna i częsta
aktywność (zjawisko desynchronizacji);
g praca cykliczna spowalnia potencjały w tempie ruchów („oznakowane rytmy” EEG); h wykonanie opanowanego wyglądu ruchowego rytmu alfa.
Medycyna i weterynaria
Neurony piramidalne. Duże komórki, perikarya o wielkości 10-130 mikronów. Komórka ma dendryt wierzchołkowy, który jest skierowany w stronę warstwy molekularnej; procesy boczne – dendryty; z podstawy wychodzi długi akson - będzie to włókno nerwu ruchowego (początek tworzenia przewodu piramidowego).
Wykład z histologii nr 13
Ośrodkowy układ nerwowy. Kora mózgowa. Cytoarchitektura warstw kory mózgowej. Skład neuronalny. Charakterystyka neuronów piramidalnych. Modułowa jednostka strukturalna i funkcjonalna kory mózgowej. Móżdżek. Struktura i skład nerwowy kory móżdżku.
Źródło rozwoju istoty szarej i białej mózgu
Rozwija się z płaszcza lub strefy płaszcza istoty szarej cewy nerwowej
Istota biała zasnówki brzeżnej
składniki istota szara mózgu (kora nowa)
6 warstw:
- Molekularny powstaje głównie w wyniku procesów zachodzących w komórkach poniżej leżących poniżej warstw, ale istnieją pojedyncze neurony, które nazywane są poziomymi.
- Zewnętrzna obecność granulatu w kształcie gwiazdy neurony i nieliczne neurony piramidowe.
- Zewnętrzna piramida (piramida) zawiera najszerszą warstwę neurony piramidalne.
- Wewnętrzny drobny granulat w kształcie gwiazdy neurony wykonują połączenia wewnątrzkorowe (nie wchodzą do istoty białej).
- Wewnętrzna piramida (zwojowa) jest reprezentowana przez dużą piramidalny neurony lub komórki Bettsa.
- Formy komórek polimorficznych są zróżnicowane, komórki są małe, a ich aksony sięgają do istoty białej.
Neurony gwiaździste.Perikarya mają okrągły lub trójkątny kształt, średnicę 4-8 mikronów; akson i dendryty tych komórek uczestniczą w połączeniach wewnątrzkorowych.
Neurony piramidalne.Duże komórki, perikarya o wielkości 10-130 mikronów. Komórka ma dendryt wierzchołkowy, który jest skierowany w stronę warstwy molekularnej; procesy boczne dendryty; długi akson pochodzi z podstawy i będzie włóknem nerwu ruchowego (początek tworzenia przewodu piramidowego). Zabezpieczenia pochodzą z aksonu: idą do sąsiednich komórek lub wznoszą się i oddziałują z leżącymi nad nimi warstwami.
ogólna charakterystyka neurony korowe:
Wszystkie neurony kory mózgowej są wielobiegunowe. Według funkcji warstwy 1-4: asocjacyjne (interkalarne), 5-6 motorowe.
Strukturalna i funkcjonalna jednostka mózgu: moduł
Moduł (w korze mózgowej do 3 milionów) pionowa kolumna o średnicy 300 mikronów, prostopadła do powierzchni kory i przechodząca przez całą grubość kory.
W środku modułu:
- Jedno włókno nerwowe korowo-stawowe jest włóknem nerwowym w korze mózgowej; pochodzące albo z neuronów piramidalnych tej samej półkuli (asocjacyjno-interkalarne), albo z półkuli przeciwnej (spoidłowe).
- Dwa włókna wzgórzowo-korowe (włókna wrażliwe na aferę), wychodzące ze wzgórza wzrokowego i biegnące w 4 warstwach; następnie powstaje odprowadzające włókno nerwowe utworzone przez aksony Najlepsze komórki.
Cytoarchitekturaobszary kory różniące się od siebie budową i umiejscowieniem komórek.
Mieloarchitekturaobszary kory różniące się między sobą charakterystyką ułożenia włókien.
Pola różne obszary kory różniące się od siebie: cytologią, mieloarchitekturą i znaczeniem funkcjonalnym.
Rodzaje kory:
- Środek wrażliwy ziarnisty, w którym dobrze rozwinięte są warstwy 2 i 4; kończą (aferentne) włókna nerwowe czuciowe wychodzące z narządów węchu, słuchu i wzroku.
- Agranularny ośrodek motoryczny, w którym rozwijają się warstwy 3, 5 i 6.
Wiarygodnym morfologicznym odpowiednikiem inteligencji jest liczba synaps neuronów asocjacyjnych (warstwy 1-4), zwykle 10 tys.
Móżdżek
Źródło rozwoju tylnej części mózgu móżdżku
Funkcje: koordynuje ruch i równowagę
Móżdżek ma dużą liczbę zwojów, w przygotowaniu w postaci rozgałęzionego drzewa.
Zakręty są oddzielone rowkami. Każdy zakręt zawiera: wąską płytkę istoty białej, całkowicie pokrytą szare komórki(kora). Skupiska neuronów leżą głęboko w istocie białej móżdżku.: jądra móżdżku.
Istota szara lub kora móżdżku ma trzy warstwy:
- Zewnętrzne molekularne są reprezentowane przez wielobiegunowe neurony asocjacyjne (interkalarne). Istnieją 2 typy komórek:
- Neurony koszykowe znajdują się w dolnej jednej trzeciej warstwy molekularnej, komórkach nieregularny kształt małe rozmiary. Akson komórki łączy się z perykareonem komórki Purkenjego, gdzie rozgałęzia się i tworzy splot w postaci kosza. Dendryty w warstwie molekularnej.
- Gwiazdy (duże i małe) są zlokalizowane na zewnątrz warstwy molekularnej (powierzchnia kory).Duże neurony gwiaździste:synapsy aksonów z perikaryą komórek Purkenjego i bierze udział w tworzeniu koszyka. Dendryty w warstwie molekularnej.Małe neurony gwiaździste: Synapsy aksonów z dendrytami komórek Purkenjego.
- Środkowy zwojowy - reprezentowany przez jedną warstwę, ciała wielobiegunowych neuronów ruchowych lub komórki Purkinjego. Są to duże komórki w kształcie gruszki z perykarionu (ciała) z 2-3 dendrytami sięgającymi do warstwy molekularnej. Akson wychodzi z ciała komórki i przechodzi przez warstwę ziarnistą do istoty białej, gdzie kończy się w jądrach móżdżku. Aksony komórek Purkinjego są jedynymi odprowadzającymi (motorycznymi) włóknami nerwowymi wychodzącymi z kory móżdżku. Bliżej ciała komórki od aksonu odchodzą zabezpieczenia, które trafiają do warstwy zwojowej i głębokich części warstwy molekularnej, gdzie łączą się z ciałami i dendrytamisąsiadujące komórki Purkiniego.
- Wewnętrzne ziarniste wszystkie komórki są polarne i skojarzone zawierają dwa typy komórek:
1-Komórki-Ziarna małe komórki, perykariony mają średnicę 4-6 mikronów, dendryty: krótkie 3-4 kawałki, skierowane w stronę istoty białej, przypominające ptasie łapki. Akson wznosi się do warstwy molekularnej, gdzie dzieli się w kształcie litery T.
2-Duża gwiaździsta: z długimi aksonami i krótkimi aksonami Komórki Golgiego drugiego typu. Duże komórki ciała są zlokalizowane bezpośrednio pod warstwą zwojową. Synapsy aksonów łączą się z rozetami włókien omszałych, zanim połączą się z dendrytami komórek ziarnistych. Dendryty wznoszą się do warstwy molekularnej.
Doprowadzające (wrażliwe) włókna nerwowe:
- Końcówka omszałych włókien włókno nerwowe nazywa się gniazdem. Synapsa rozet omszonych włókien łączy się z dendrytami komórek ziarnistych, tworząc kłębuszki móżdżku.
- Włókna pnące lub liany pochodzą z istoty białej, przechodzą przez warstwy ziarniste i łączą się z perykarionem komórki Purkinjego lub z dendrytami tych komórek.
Aksony synaps komórek ziarnistych z dendrytami:
- Warstwa molekularna komórki
- Komórki Purkenjego
- Komórka Golgiego typu II
Układ hamujący móżdżku obejmuje:
- Komórki warstwy molekularnej
- Komórki Golgiego typu 2
- Synapsy pobudzające:
- Włókna omszałe
- Włókna Liany
Jak również inne prace, które mogą Cię zainteresować |
|||
| 73026. | Wprowadzanie, formatowanie danych i pisanie formuł | 126KB | |
| Cel: Użyj polecenia Formatuj komórki, aby sformatować dane w kolumnie D w formacie walutowym bez miejsc dziesiętnych. Za pomocą przycisku Wzorzec formatu skopiuj format kolumny D do E. Użyj przycisków na panelu Formatowanie, aby określić format procentowy w kolumnie F z dwoma miejscami po przecinku. | |||
| 73027. | Tworzenie i edycja wykresów w programie Excel | 160KB | |
| Naucz się tworzyć diagramy za pomocą kreatora; Naucz się edytować diagramy. Pytania testowe Do czego służą diagramy. Jakie znasz rodzaje diagramów. Jak zbudować diagram osobny arkusz Jak zmienić typ wykresu Jak usunąć wykres... | |||
| 73028. | Modelowanie systemów plików | 147,5 kB | |
| Użytkownicy nadają plikom symboliczne nazwy, biorąc pod uwagę pewne ograniczenia systemu operacyjnego. Katalog zawiera listę zawartych w nim plików i ustala zgodność pomiędzy plikami oraz ich charakterystykami i atrybutami. | |||
| 73029. | Wyznaczanie składu strukturalno-fazowego NVM zastępującego VNT metodami dyfrakcji rentgenowskiej i mikroskopii elektronowej | 1,5 MB | |
| Istotna jest struktura fazowo-strukturalna NVM, aby dopasować VNT do danych z dyfrakcji rentgenowskiej i mikroskopii elektronowej. Obserwuj zmianę składu faz strukturalnych NVM podczas obróbki termochemicznej. | |||
| 73033. | Niestandardowe typy danych na przykładzie MS SQL Server | 51KB | |
| Poznaj możliwości definiowania i używania niestandardowych typów danych, wyzwalaczy DML i DDL oraz ograniczeń integralności w postrelacyjnym systemie DBMS. Opanuj metody i technologie tworzenia niestandardowych typów danych w CLR.Net. | |||
| 73034. | Zasada oznakowania żelbetowych płyt stropowych budynków cywilnych | 47,1 kB | |
| Zasada znakowania żelbetowych płyt stropowych budynków cywilnych Zgodnie z GOST 2300978 płyty stropowe są oznaczane szeregiem indeksów alfanumerycznych. Pierwsza grupa zawiera oznaczenie rodzaju płyty oraz jej wymiary konstrukcyjne, długość i szerokość w decymetrach. | |||
System nerwowy. Wykład 2.
Kora Kora mózgowa (cortex cerebri) to ciągła warstwa istoty szarej na powierzchni półkul mózgowych o grubości 2-5 mm. Znajduje się nie tylko na zwojach, ale przechodzi we wszystkie bruzdy, przez co jego powierzchnia jest stosunkowo duża i u dorosłego osobnika wynosi 2200 cm 2 . Masa kory wynosi 581 g, jej objętość wynosi około 560 cm3. W tym przypadku główna objętość kory składa się z istoty białej (450 cm 3, waga - 470 g). Masa ciała komórkowego neuronów korowych wynosi tylko około 21 g (20 cm3).
Rozmiar korowych komórek nerwowych waha się w szerokim zakresie od 8-9 mk do 150 mk. W korze mózgowej człowieka znajduje się aż 15 miliardów komórek nerwowych, z czego 6 miliardów to komórki małe. Zdecydowana większość neuronów korowych należy do dwóch typów: neuronów piramidalnych i neuronów gwiaździstych. Komórki te znajdują się w korze mózgowej w określonych warstwach.
W mózgu człowieka wyróżnia się kilka filogenetycznie różne rodzaje kora:
Starożytna kora - paleocortex (0,6%) - praktycznie nie ma warstw (1-2 warstwy), jest zlokalizowana w obszarze trójkątów węchowych, tworzy osłonę przegrody przezroczystej i otacza jądro ciała migdałowatego.
Stara kora - archicortex (2,2%) - ma 2-3 warstwy, reprezentowane przez istotę szarą zakrętu zębatego, istotę szarą hipokampa i prążki rdzeniowe na ciele modzelowatym.
Kora przejściowa - mezokora (1,6%) - obszar sklepionego zakrętu.
Nowa kora – kora nowa (95,6%) – jest dobrze zbudowana i posiada 6 warstw.
U wyższych kręgowców występuje przeważnie sześć mniej lub bardziej wyraźnie określonych warstw. Ale każdą z tych warstw, z wyjątkiem pierwszej, można podzielić na dwie lub nawet trzy podwarstwy.
Pierwsza warstwa, tak zwany strefowy lub płytka molekularna, składa się głównie ze splotów wierzchołkowych dendrytów neuronów piramidowych, których ciała komórkowe znajdują się w innych warstwach kory. W pierwszej warstwie jest bardzo mało komórek nerwowych. Zawiera komórki poziome, których aksony i dendryty są również umieszczone poziomo w tej samej warstwie.
Druga warstwa, tzw. Zewnętrzny granulat lub zewnętrzna płyta ziarnista, obejmuje masę małych komórek należących do małych piramidalnych, tak zwanych interneuronów i neuronów gwiaździstych, przy czym w drugiej warstwie dominują małe komórki piramidalne.
Trzecia warstwa to zewnętrzna płyta piramidalna zawiera średniej wielkości komórki piramidalne
Czwarta warstwa, tak zwany wewnętrzny granulat lub wewnętrzna płyta ziarnista, składa się głównie z małych komórek gwiaździstych, ale zawiera także małe i średnie piramidy.
Piąta warstwa to wewnętrzna płyta piramidalna zawiera gigantyczne komórki piramidalne lub komórki Betza.
Szósta warstwa – płyta wielopostaciowa zawierają głównie średniej wielkości komórki piramidalne i niewielką liczbę małych komórek piramidalnych i gwiaździstych.
Pierwsze trzy warstwy są najmłodsze i zapewniają połączenie pomiędzy różnymi odcinkami kory. Czwarta warstwa jest najbardziej rozwinięta w obszarach, do których dociera informacja doprowadzająca (ośrodki wrażliwe, zwłaszcza zakręt postcentralny). Piąta warstwa wyraża się w obszarach motorycznych kory: zakręt przedśrodkowy, płatek okołośrodkowy, zakręt nadbrzeżny.
Każdy neuron piramidalny ma komórkę piramidalną i wiele dendrytów. Akson neuronu piramidalnego wyłania się z małego wzgórka u podstawy komórki. W małych, tak zwanych neuronach interkalarnych lub pośrednich piramidalnych, aksony rozgałęziające się w kierunku poziomym lub pionowym kończą się natychmiast, nie opuszczając kory. Aksony średnich i dużych piramid wydzielają wiele zabezpieczeń w korze, a główne pnie przechodzą do podkorowej istoty białej. Część z nich wraca z substancji podkorowej do kory danej półkuli lub przechodząc przez ciało modzelowate kończy się w korze drugiej półkuli. Służą do łączenia różnych części kory mózgowej. Dlatego nazywane są takie neurony piramidalne asocjacyjny. Pozostałe aksony kierowane są do formacji podkorowych i dalej do różnych części mózgu i rdzenia kręgowego. Te piramidy nazywają się występ. W związku z rozwojem filogenetycznym ssaków liczba komórek piramidalnych znacznie wzrasta.
Szerokość całej kory, liczba komórek i szerokość każdej warstwy komórek w części czołowej, a także skład komórkowy, tj. Rozmiar, kształt i rozmieszczenie komórek w każdej części kory są bardzo zróżnicowane . Odmiany te zajmują pewne, wyraźnie wyznaczone terytoria. W mózgu dużego ssaka znajduje się 11 wyraźnie widocznych dużych pól korowych.
Na podstawie badań histologicznych i fizjologicznych każde takie pole podzielono na kilka niezależnych sekcji. Większe zróżnicowanie kory obserwuje się u wyższych ssaków – małp. Jest to szczególnie wspaniałe u ludzi.
Badania takie rozpoczął kijowski naukowiec Władimir Aleksiejewicz Bets, który w 1874 roku opublikował artykuł „Dwa ośrodki w korze mózgowej”, w którym opisał strefę motoryczną w zakręcie przedśrodkowym i strefę wrażliwą w zakręcie postcentralnym.
W 1909 roku niemiecki neurolog Korbinian Brodmann opublikował mapy pól cytoarchitektonicznych Kora mózgowa. Brodmann jako pierwszy stworzył mapy kory mózgowej. Następnie O. Vogt i C. Vogt (1919-1920), biorąc pod uwagę strukturę włókien, opisali 150 obszarów mieloarchitektonicznych w korze mózgowej. W Instytucie Mózgu Akademii Nauk Medycznych ZSRR I. N. Filippov i S. A. Sarkisov stworzyli mapy kory mózgowej obejmujące 47 pól cytoarchitektonicznych.
Dane badania eksperymentalne wskazują, że zniszczenie lub usunięcie pewnych obszarów kory mózgowej u zwierząt powoduje zakłócenie niektórych funkcji. Fakty te zostały potwierdzone obserwacje kliniczne dla chorych ze zmianami nowotworowymi lub urazami niektórych obszarów kory mózgowej. Wszystko to pozwoliło nam stwierdzić, że w korze mózgowej znajdują się ośrodki regulujące wykonywanie określonych funkcji. Morfologicznym potwierdzeniem danych fizjologicznych i klinicznych była doktryna o różnej jakości struktury kory mózgowej w różnych jej częściach - cyto- i mieloarchitektoniki kory. Ustalono, że neurony nie są rozmieszczone rozproszonie w korze, ale są pogrupowane w zespoły.
Zastosowanie nowoczesnych metod mikroelektrodowych do badania funkcji neuronów korowych znacznie poszerzyło wiedzę na temat przetwarzania informacji sensorycznej w korze nowej i organizacji strukturalnej kory. W 1957 roku amerykański badacz V. Mountcastle, analizując reakcje komórek kory somatosensorycznej (czuciowo-ruchowej) kota na bodźce o różnej modalności, odkrył co następuje: interesujący fakt. Kiedy mikroelektrodę zanurzono prostopadle do powierzchni kory somatosensorycznej, wszystkie napotkane komórki reagowały jednakowo na bodziec, na przykład na lekki dotyk skóry lub ruch w stawie. Jeśli elektroda została zanurzona pod kątem do powierzchni kory, wówczas wzdłuż jej ścieżki pojawiały się neurony o różnych modalnościach sensorycznych, na przemian z pewną okresowością.
Na podstawie tych eksperymentalnych faktów W. Mountcastle doszedł do wniosku, że kora somatosensoryczna jest zorganizowana w elementarne jednostki funkcjonalne - kolumny zorientowane prostopadle do powierzchni. Średnica takiej kolumny wynosi około 500 μm, co wynika z poziomego rozmieszczenia końcówek doprowadzającego włókna wzgórzowo-korowego i pionowej orientacji dendrytów komórki piramidalne. Według Mountcastle kolumna tak elementarny blok kory sensomotorycznej, gdzie odbywa się lokalne przetwarzanie informacji z receptorów jednej modalności. Ta hipoteza o kolumnowej organizacji kory nowej stała się powszechna i dała impuls do dalszych badań w tym zakresie w kraju i za granicą.
Według nowoczesne pomysły, każda kolumna funkcjonalna kory sensomotorycznej składa się z kilku mikromodułów morfologicznych, które łączą od pięciu do sześciu neuronów przypominających gniazda. Moduł ten zawiera kilka komórek piramidalnych, których wierzchołkowe dendryty są jak najbliżej siebie i tworzą wiązkę dendrytyczną; w obrębie tej wiązki są możliwe połączenia elektrotoniczne, które najprawdopodobniej zapewniają synchroniczne działanie całego stowarzyszenia.
Do grupy pionowo zorientowanych komórek piramidalnych przylegają komórki gwiaździste, z którymi stykają się włókna wzgórzowo-korowe dochodzące do mikromodułu. Niektóre komórki gwiaździste pełniące funkcję hamującą mają długie aksony rozciągające się poziomo. Aksony komórek piramidalnych tworzą powtarzające się zabezpieczenia, które mogą zapewniać zarówno wpływy ułatwiające w obrębie mikromodułu, jak i interakcje hamujące między mikromodułami, kontaktując się z interneuronami hamującymi. Kilka mikromodułów strukturalnych, połączonych poziomym rozgałęzieniem zakończeń określonych włókien doprowadzających wzgórzowo-korowych, aksonów zakończeń i procesów komórek gwiaździstych, tworzy kolumnę (lub makromoduł), którego średnica sięga 500-1000 µm. Każda kolumna charakteryzuje się jednością funkcjonalną, która objawia się tym, że neurony kolumny odpowiadają na bodziec jednej modalności.
Następnie zasada organizacji kolumnowej została potwierdzona podczas badania innych strefy projekcyjne kora.
IP Pawłow uważał korę mózgową za ciągłą powierzchnię percepcyjną, za zbiór korowych końcówek analizatorów. Pokazał, że koniec korowy analizatorów nie jest jakąś ściśle określoną strefą. Kora mózgowa jest podzielona na jądro i rozproszone elementy. Jądro jest miejscem koncentracji komórek nerwowych kory mózgowej, stanowiącym dokładną projekcję wszystkich elementów specyficznego receptora obwodowego, gdzie zachodzi wyższa analiza, synteza i integracja funkcji. Elementy rozproszone mogą być zlokalizowane zarówno wzdłuż obwodu jądra, jak i w znacznej odległości od niego. Dokonują prostszej analizy i syntezy. Obecność rozproszonych elementów podczas niszczenia rdzenia pozwala częściowo zrekompensować upośledzoną funkcję. Obszary zajmowane przez rozproszone elementy różne analizatory, można układać warstwami jedna na drugiej, zachodzić na siebie. Zatem korę mózgową można schematycznie wyobrazić jako zbiór jąder różnych analizatorów, pomiędzy którymi rozproszone są elementy należące do różnych (sąsiadujących ze sobą) analizatorów. Powyższe pozwala nam o tym porozmawiać dynamiczna lokalizacja funkcji w korze mózgowej (I.P. Pavlov).
Rozważmy położenie niektórych końców korowych różnych analizatorów (jąder) w stosunku do zakrętów i płatów półkul mózgowych u człowieka zgodnie z mapami cytoarchitektonicznymi.
1. W korze zakrętu postśrodkowego i płatka ciemieniowego górnego znajdują się komórki nerwowe, które tworzą rdzeń analizatora korowego ogólnego wrażliwość(temperatura, ból, dotyk) i proprioceptywne. Drogi czuciowe prowadzące do kory mózgowej przecinają się albo na poziomie różnych odcinków rdzenia kręgowego (ścieżki bólu, wrażliwości na temperaturę, dotyk i ucisk), albo na poziomie rdzenia przedłużonego (ścieżki wrażliwości proprioceptywnej kierunku korowego ). W rezultacie zakręty postcentralne każdej półkuli są połączone z przeciwną połową ciała. W zakręcie postcentralnym wszystkie pola receptorowe różnych części ciała ludzkiego są rzutowane w taki sposób, że korowe końce analizatora wrażliwości dolnych partii ciała i dolne kończyny, a pola receptorowe górnych partii ciała i głowy oraz kończyn górnych są wystające najniżej (bliżej bruzdy bocznej).
2. Rdzeń silnik analizator zlokalizowany jest głównie w tzw. korze ruchowej, która obejmuje zakręt przedśrodkowy i płatek paracentralny na powierzchnia środkowa półkule. W piątej warstwie kory zakrętu przedśrodkowego leżą neurony piramidalne (komórki Betza), które I.P. Pavlov zaklasyfikował jako interkalarne i zauważył, że komórki te swoimi procesami są połączone z jądrami podkorowymi, komórkami motorycznymi jąder czaszki i nerwy rdzeniowe. Ponadto w górnych partiach zakrętu przedśrodkowego i w płacie przyśrodkowym znajdują się komórki, z których impulsy przekazywane są do mięśni najniższych partii tułowia i kończyn dolnych. W dolnej części zakrętu przedśrodkowego znajdują się ośrodki motoryczne regulujące aktywność mięśni twarzy.
W ten sposób wszystkie obszary ludzkiego ciała są rzutowane „do góry nogami” w zakręcie przedśrodkowym. Ze względu na to, że drogi piramidowe, wywodzące się z komórek gigantopiramidalnych, przecinają się albo na poziomie pnia mózgu (włókna korowo-jądrowe), na granicy z rdzeniem kręgowym, albo w odcinkach rdzenia kręgowego (droga korowo-rdzeniowa), motoryka obszary każdej półkuli są połączone z mięśniami szkieletowymi przeciwnej strony ciała. Jeśli mięśnie kończyn są połączone izolowanie z jedną z półkul, mięśnie tułowia, krtani i gardła są połączone z obszarami motorycznymi obu półkul.
Silnik I.P. Pavlov nazwał także obszar kory mózgowej obszarem receptorowym, ponieważ tutaj analiza proprioceptywnych (bodźców kinetycznych odbieranych przez receptory osadzone w mięśnie szkieletowe, ścięgna, powięzi i torebki stawowe).
3. Rdzeń analizatora realizujący tę funkcję połączony obrót głowy i oczu przeciwna strona , jest w rejony tylneśrodkowy zakręt czołowy, w tzw. strefie przedruchowej.
4. W obszarze płatka ciemieniowego dolnego, w zakręcie nadbrzeżnym, znajduje się jądro analizatora motorycznego, którego znaczenie funkcjonalne polega na realizacji synteza wszyscy ruchy celowe, zawodowe, robotnicze i sportowe. Rdzeń ten jest asymetryczny. Dla praworęcznych jest po lewej stronie, a dla leworęcznych- tylko w prawej półkuli. Zdolność do koordynowania tych złożonych, celowych ruchów jednostka nabywa przez całe życie w wyniku praktycznej aktywności i gromadzenia doświadczeń. Realizacja ukierunkowanych ruchów następuje w wyniku tworzenia tymczasowych połączeń między komórkami zlokalizowanymi w zakrętach przedśrodkowych i nadbrzeżnych. Uszkodzenie pola nie powoduje paraliżu, a jedynie prowadzi do utraty zdolności do wykonywania złożonych, skoordynowanych, celowych ruchów - apraksja (praksja - praktyka).
5. W korze płatka ciemieniowego górnego znajduje się jądro analizatora skóry o jednym ze szczególnych typów wrażliwości, które pełni nieodłączną funkcję rozpoznawanie obiektów poprzez dotyk,- streognozja. Korowa końcówka tego analizatora znajduje się w prawej półkuli. Uszkodzeniu powierzchniowych warstw kory w tym odcinku towarzyszy utrata funkcji rozpoznawania obiektów za pomocą dotyku, chociaż zachowane są inne rodzaje ogólnej wrażliwości.
6. W głębi bruzdy bocznej, na powierzchni środkowej części zakrętu skroniowego górnego zwróconego w stronę wyspy (gdzie widoczne są zakręty poprzeczne skroniowe, czyli zakręty Heschla) znajduje się jądro słuchowy analizator. DO komórki nerwowe, stanowiący rdzeń analizator słuchowy każda z półkul, ścieżki przewodzące z receptorów zarówno lewej, jak i prawa strona. Pod tym względem jednostronne uszkodzenie tego jądra nie powoduje całkowitej utraty zdolności odbierania dźwięków. Obustronnemu uszkodzeniu towarzyszy głuchota korowa.
7. Rdzeń wizualny analizator znajduje się w płata potylicznego półkule mózgowe na obwodzie bruzdy kalkarynowej. Rdzeń analizator wizualny prawa półkula jest połączona ścieżkami z boczną połową siatkówki prawego oka i przyśrodkową połową siatkówki lewego oka. W korze płata potylicznego lewej półkuli rzutowane są odpowiednio receptory bocznej połowy siatkówki lewego oka i środkowej połowy siatkówki prawego oka. Podobnie jak w przypadku jądra analizatora słuchowego, jedynie obustronne uszkodzenie jąder analizatora wzrokowego prowadzi do całkowitej ślepoty korowej. Uszkodzeniu pola znajdującego się nieco powyżej jądra analizatora wizualnego towarzyszy utrata pamięci wzrokowej, ale nie obserwuje się utraty wzroku. Jeszcze wyżej znajduje się część kory, której porażce towarzyszy utrata zdolności poruszania się w nieznanym środowisku.
8. Na dolnej powierzchni płata skroniowego półkuli mózgowej, w obszarze uncus i częściowo w obszarze hipokampu znajduje się jądro węchowy analizator. Z filogenetycznego punktu widzenia obszary te należą do najstarszych części kory mózgowej. Zmysły węchu i smaku są ze sobą ściśle powiązane, co tłumaczy się bliskim położeniem jąder analizatorów węchowych i smakowych. Zauważono również (V.M. Bekhterev), że percepcja smaku jest zaburzona, gdy uszkodzona jest kora najniższych części zakrętu postcentralnego. Jądra analizatorów smaku i węchu obu półkul są połączone z receptorami zarówno po lewej, jak i prawej stronie ciała.
Opisane końce korowe niektórych analizatorów nie występują w korze mózgowej tylko ludzie, ale także zwierzęta. Specjalizują się w percepcji, analizie i syntezie sygnałów pochodzących ze środowiska zewnętrznego i wewnętrznego, stanowiących w rozumieniu I.P. Pavlova pierwszy system sygnałowy rzeczywistości.
Drugi system sygnalizacji istnieje tylko u ludzi i jest zdeterminowany rozwojem mowy. Mowa, a wraz z nią świadomość, są filogenetycznie najmłodszymi funkcjami mózgu. Pod tym względem korowe końce analizatorów są najmniej zlokalizowane. Funkcje mowy i myślenia realizowane są przy udziale całej kory mózgowej. Jednak w korze mózgowej można wyróżnić pewne obszary, które charakteryzują się ściśle określonymi funkcjami mowy. Tak, analizatory mowa motoryczna(ustne i pisemne) znajdują się obok obszaru motorycznego kory, a dokładniej w tych częściach kory płata czołowego, które sąsiadują z zakrętem przedśrodkowym.
Analizatory wizualne i percepcja słuchowa sygnały mowy zlokalizowane są obok analizatorów wzroku i słuchu. Należy podkreślić, że analizatory mowy u osób praworęcznych zlokalizowane są w lewej półkuli, a u osób leworęcznych – głównie w prawej.
Rozważmy położenie niektórych analizatorów mowy w korze mózgowej.
1. Analizator motoryczny mowy pisanej znajduje się w tylnej części środkowego zakrętu czołowego. Działanie tego analizatora jest ściśle powiązane z analizatorem celowych ruchów, który zapewnia kształtowanie wyuczonych ruchów dłoni i oczu niezbędnych do pisania. W przypadku uszkodzenia traci się zdolność wykonywania drobnych ruchów, zwykle wykonywanych pod kontrolą wzroku i niezbędnych do rysowania liter, słów i innych znaków (agrafia).
2. Rdzeń analizatora silnika artykulacja mowy(analizator silnika mowy) znajduje się w tylnych częściach dolnego zakrętu czołowego (centrum Broki). Uszkodzenie kory w tym obszarze prowadzi do afazji ruchowej, czyli utraty zdolności wymawiania słów (afazji). Ta afazja nie jest związana z utratą zdolności do kurczenia się mięśni zaangażowanych w wytwarzanie mowy. Co więcej, jeśli to pole zostanie uszkodzone, zdolność wymawiania dźwięków lub śpiewania nie zostanie utracona.
3. B wydziały centralne znajduje się dolny zakręt czołowy rdzeń analizatora mowy związany ze śpiewem. Jego porażce towarzyszy amuzja wokalna – niemożność komponowania i odtwarzania fraz muzycznych oraz agramatyzm, gdy zatraca się umiejętność komponowania sensownych zdań z poszczególnych słów. Mowa takich ludzi składa się z zestawu słów, które nie są ze sobą powiązane w znaczeniu semantycznym.
4. Rdzeń analizator mowy słuchowej blisko powiązane z
centrum korowe analizatora słuchowego i znajduje się w obszarach tylnych
zakręt skroniowy górny, na swojej powierzchni skierowany w bok
bruzda boczna półkuli mózgowej. Uszkodzenie jądra nie upośledza ogólnie słuchowego postrzegania dźwięków, ale następuje utrata zdolności rozumienia słów i mowy (głuchota werbalna lub afazja sensoryczna). Funkcją tego rdzenia jest to, że osoba nie tylko słyszy i rozumie mowę innej osoby, ale także kontroluje swoją.
5. W środkowej jednej trzeciej górnego zakrętu skroniowego znajduje się jądro analizatora korowego, którego porażce towarzyszy początek głuchoty muzycznej, gdy frazy muzyczne są postrzegane jako bezsensowny zestaw różnych dźwięków. Ten korowy koniec analizatora słuchowego należy do ośrodków drugiego układu sygnałowego, które postrzegają werbalne oznaczenie obiektów, działań, zjawisk, tj. sygnały wykrywające sygnał.
6. W bezpośrednim połączeniu z rdzeniem analizatora wizualnego znajduje się rdzeń wizualny analizator mowy pisanej(pole 39), zlokalizowane w zakręcie kątowym płatka ciemieniowego dolnego. Uszkodzenie tego jądra prowadzi do utraty zdolności postrzegania tekstu pisanego i czytania (dysleksja).
Ludzka kora czołowa jest powiązana z zasadą społeczną. Jego neurony tworzą dwukierunkowe połączenia ze wszystkimi innymi obszarami kory. Wraz z uszkodzeniem powstają zaburzenia zachowanie społeczne, chamstwo, nielojalność, obniżone poczucie celu, apatia. Po lobotomii następuje całkowity brak inicjatywy i aktywności społecznej.
Asymetria funkcji prawej i lewej półkuli jest bardzo wyraźna. Lewa półkula zapewnia logikę, abstrakcyjne myślenie, dominuje w odniesieniu do funkcji mowy (czytanie, pisanie, liczenie) i złożonych ruchów dowolnych. Prawa półkula– emocjonalność, myślenie wyobraźniowe, pozamowe funkcje rozpoznawania złożonych obrazów wzrokowych i słuchowych, percepcja dotykowa, postrzeganie przestrzeni, kształtu, kierunku, myślenie intuicyjne.
Podobne artykuły
-
Sochni z mąki żytniej Sochni na Wniebowstąpienie
Sochen to placek złożony na pół z nadzieniem. Osobliwością sochnii (w przeciwieństwie do prawdziwych ciast) jest to, że nie jest ona ściskana i że ciasto drożdżowe nie wyrasta i nie wychodzi, ale jest krojone i natychmiast wkładane do piekarnika. Dlatego...
-
Żyto sochni z twarogiem. Sok z mąki żytniej. Sochni o Wniebowstąpienie
Pomysł na soki żytnie zaczerpnąłem od mike_cooking, który natknął się na ten cud podczas wyprawy etno-kulinarnej. Przepis wybrałam w oparciu o przepis na „zwykłe” soki pszenne i instynktownie :) Pokhlebkin twierdzi jednak, że sok będziemy robić na...
-
Kompot jabłkowy na zimę - niedrogie przepisy w domu
Przepisy krok po kroku na kompot jabłkowy na zimę: klasyczny, szybki i łatwy w powolnym naczyniu bez cukru, niebiański kompot z miętą, agrestem, wiśniami, winogronami 2018-06-14 Irina Naumova Ocena przepisu 846...
-
Napój deserowy - galaretka skrobiowa
To zależy od tego, jak gęstą masz galaretkę. A także - na jakość skrobi. Czasami skrobia jest kiepskiej jakości - nie daje dobrej konsystencji, niezależnie od ilości jej dodanej. Zwykle na opakowaniu jest napisane, ile łyżek skrobi potrzebujesz...
-
Jak zamrozić arbuza w domu: proste przepisy na przygotowanie go na zimę Czy można jeść mrożonego arbuza?
Arbuz to duża, słodka jagoda, którą lubi wielu ludzi. Niestety okres, w którym można cieszyć się nim z głębi serca, jest krótki, ale za to bardzo miło jest zjeść kawałek soczystego miąższu arbuza w chłodny jesienny wieczór lub ugotować pyszną...
-
Sałatka noworoczna bez majonezu
Prawdziwa magia i niespokojne oczekiwanie na cud, migotanie świec i blask bujnego blichtru, zimowa zabawa, długo oczekiwane prezenty i świąteczna uczta - wszystko to czeka na nas w przeddzień Nowego Roku 2017. Jeśli prezenty dla przyjaciół i rodziny są już...