A tudósok megszámolták az emberi agyban lévő neuronok számát. A kortikális aktivitás elektromos megnyilvánulásai
A féltekék kérge nagy agy a legmagasabb és legösszetettebb szervezetet képviseli idegközpont képernyőtípus, melynek tevékenysége biztosítja a különböző testfunkciók és komplex magatartásformák szabályozását.
A kéreg 3-5 mm vastag szürkeállományból áll. A szürkeállomány idegsejteket (több mint 10 milliárd neuront), idegrostokat és neurogliasejteket tartalmaz. Különböző szakaszait, amelyek a sejtek elhelyezkedésének és szerkezetének, a rostok elhelyezkedésének és funkcionális jelentőségének bizonyos jellemzőiben különböznek egymástól, mezőknek nevezik, amelyeket először K. Brodmann német orvos és tudós írt le.
Citoarchitektúra
A kéreg multipoláris neuronjai közül piramis-, csillag-, orsó alakú, pókszerű, vízszintes, „kandeláber”-sejtek, kettős dendritcsokorral rendelkező sejtek és néhány más típusú neuron különböztethető meg.
Piramis neuronok az agykéreg alapvető és legspecifikusabb formáját alkotják. Hosszúkás kúp alakú testük van, amelynek csúcsa a kéreg felülete felé néz. A dendritek a test csúcsáról és oldalsó felületeiről nyúlnak ki. Az axonok a piramissejtek alapjából származnak.
A kéreg különböző rétegeinek piramissejtek mérete különbözik és eltérő funkcionális érték. A kis sejtek interneuronok. A kialakulásban nagy piramisok axonjai vesznek részt motor piramis utak.
A kéreg neuronjai homályosan körülhatárolt rétegekben helyezkednek el, amelyeket római számokkal jelölnek és kívülről befelé számoznak. Minden réteget egy-egy sejttípus túlsúlya jellemez. Az agykéregben hat fő réteg van:
- I - molekuláris;
- II - külső szemcsés;
- III - piramis alakú;
- IV - belső szemcsés;
- V - ganglionos;
- VI - polimorf sejtek rétege.
én - Molekuláris a kéregréteg nem tartalmaz nagyszámú a Cajal kis asszociatív vízszintes sejtjei. Axonjaik párhuzamosan futnak az agy felszínével a molekuláris réteg idegrostjaiból álló tangenciális plexus részeként. Ennek a plexusnak a rostjainak nagy részét azonban az alatta lévő rétegek dendriteinek elágazása jelenti.
II - Külső szemcsés a réteget számos kis piramis- és csillagneuron alkotja. Ezeknek a sejteknek a dendritjei a molekuláris rétegbe emelkednek, és az axonok vagy bemennek fehér anyag, vagy íveket képezve a molekularéteg tangenciális rostfonatába is belépnek.
III - Az agykéreg legszélesebb rétege - piramis alakú. Piramis neuronokat, Martinotti sejteket és orsósejteket tartalmaz dupla dendritcsokorral. A piramisok csúcsi dendritjei benyúlnak a molekuláris rétegbe, és az oldalsó dendritek szinapszisokat alkotnak ennek a rétegnek a szomszédos sejtjeivel. A piramissejtek axonja mindig az alapjától nyúlik ki. Kis sejtekben a kéregben marad, a nagy sejtekben mielinrostot képez, amely az agy fehérállományába kerül. A kis poligonális Martinotti-sejtek axonjai a molekuláris rétegbe irányulnak. A piramisréteg elsősorban asszociatív funkciókat lát el.
IV - Belső szemcsés a kéreg egyes területein a réteg nagyon fejlett (például a kéreg vizuális és hallási területein), míg másokban szinte hiányozhat (például a precentralis gyrusban). Ezt a réteget kis csillagneuronok alkotják. Nagyszámú vízszintes szálat tartalmaz.
V- Ganglionos a kéreg rétegét nagy piramisok alkotják, és a motoros kéreg területe (precentrális gyrus) óriás piramisok , amelyeket először V. A. Bets kijevi anatómus írt le. A piramisok csúcsi dendritjei elérik az első réteget. A piramisok axonjai az agy motoros magjaira vetülnek és gerincvelő. A piramispályákban a Betz-sejtek leghosszabb axonjai elérik a gerincvelő caudalis szegmenseit.
A kéreg ganglionrétege a piramis neuronokon kívül függőleges orsó alakú sejteket tartalmaz, amelyek axonjai a 1. réteg kéreg, valamint kosársejtek.
VI - Réteg polimorf sejtek különböző alakú neuronok (fusiform, stellate, Martinotti sejtek) alkotják. Ezen sejtek axonjai az efferens útvonalak részeként benyúlnak a fehérállományba, és a dendritek elérik a molekuláris réteget.
Mieloarchitektúra
Az agykéreg idegrostjai között megkülönböztethetünk asszociációs az egyik félteke kéregének egyes területeit összekötő rostok, komisz, a különböző agyféltekék kéregét összekötő, és kivetítés afferens és efferens rostok, amelyek összekötik a kéreget a központi idegrendszer alsó részeinek magjaival. Az agykéregben lévő projekciós rostok radiális sugarakat képeznek, amelyek a harmadik piramisrétegben végződnek. Az I - molekuláris réteg már ismertetett tangenciális plexusa mellett a IV - belső szemcsés és V - ganglionréteg szintjén két tangenciális myelin idegrostréteg található - rendre a Baillarger külső csíkja és a belső csík. Baillarger. Az utolsó két rendszer az afferens szálak terminális szakaszaiból kialakított plexus.
A kéreg szerveződésének moduláris elve
Az agykéregben neuronok ismétlődő blokkjait vagy moduljait írják le, amelyeket annak viszonylag autonóm tevékenységre képes morfofunkcionális egységeinek tekintenek. Henger alakúak vagy oszlopok, amelyek függőlegesen futnak át a kéreg teljes vastagságán.
Minden modul tartalmaz afferens útvonalakat, helyi kapcsolatok rendszerét és efferens útvonalakat.
NAK NEK afferens pályák ide tartoznak a cortico-corticalis és a thalamo-corticalis rostok.
A modul cortico-corticalis rostok köré szerveződik, amelyek egyazon vagy az ellenkező féltekén lévő piramissejtek axonjai. A cortico-corticalis rostok egy adott modul kéregének minden rétegében végződéseket képeznek.
A modul tartalmaz továbbá thalamo-kortikális rostokat, amelyek a kéreg IV rétegében végződnek tüskés stellate neuronokon és a piramis neuronok bazális dendritjein.
Efferens utak A nagy és óriás piramis neuronok axonjai, valamint a fusiform axonok és a kéreg VI. rétegének néhány más sejtje alkotja.
Helyi kapcsolatrendszer a modul interneuronjai alkotják, amelyek több mint egy tucat sejttípust tartalmaznak. Legtöbbjük gátló hatású, és szabályozza a túlnyomórészt piramis alakú neuronok aktivitását.
A modul gátló neuronjaiból legmagasabb érték rendelkezik:
- axo-axonális sejtek;
- "kandeláber" sejtek;
- kosárcellák;
- sejtek dupla dendritcsokorral;
- sejteket axonkefével.
A gátló neuronok rendszere szűrő szerepet tölt be, gátolva a kéreg piramis neuronjainak egy részét.
Az agy és a gerincvelő agyhártyája
Az agyat és a gerincvelőt három kötőszöveti membrán borítja: puha, közvetlenül az agyszövettel szomszédos, arachnoid és kemény, amely határos. csontszövet koponya és gerinc. Az agyhártya védelmet nyújt, pl. lengéscsillapító funkció, biztosítja a cerebrospinális folyadék termelését és felszívódását.
Pia mater közvetlenül az agyszövet mellett, és attól a marginális gliahártya határolja el. Laza rostosban kötőszöveti A héj nagyszámú véredényt tartalmaz, amelyek ellátják az agyat, számos idegrostot, terminális berendezést és egyetlen idegsejtet. A pia mater körülveszi az agyba behatoló ereket, körülöttük perivaszkuláris piálmembránt képezve. Az agy kamráiban a pia mater az ependymával együtt részt vesz a cerebrospinális folyadékot termelő choroid plexusok kialakításában.
Pókhálószerű laza rostos kötőszövet vékony rétege képviseli. Közte és a pia mater között vékony kollagénkötegekből és vékony rugalmas rostokból álló keresztrudak hálózata található. Ez a hálózat köti össze a héjakat egymással. A pia mater, amely követi az agyszövet domborulatát, és az arachnoidus között, amely megemelkedett területeken fut anélkül, hogy bemélyedne, van egy subarachnoidális (subarachnoidális) tér, amelyet vékony kollagén és rugalmas rostok átjárnak, amelyek összekötik a membránokat egymással. Egyéb. A subarachnoidális tér kommunikál az agy kamráival, és cerebrospinális folyadékot tartalmaz. Ezen a téren nagyok véredény, melynek ágai táplálják az agyat.
Az arachnoid membrán bolyhjai (a legnagyobbakat Pachion granulátumnak nevezik) olyan helyként szolgálnak, amelyen keresztül az agy-gerincvelői folyadékból származó anyagok visszatérnek a vérbe. Ezek az arachnoid membrán vaszkuláris kinövései, amelyek résszerű terek hálózatát tartalmazzák, és a dura mater sinusainak lumenébe nyúlnak be.
Dura mater sok rugalmas rostot tartalmazó sűrű rostos kötőszövet alkotja. A koponyaüregben szorosan összenőtt a periosteummal. BAN BEN gerinccsatorna a dura matert a csigolya periosteumtól az epidurális tér határolja, laza rostos kötőszövetréteggel töltve, amely némi mozgékonyságot biztosít számára. A dura mater és az arachnoid membrán között van a szubdurális tér. A szubdurális tér kis mennyiségű folyadékot tartalmaz.
A szubdurális és subarachnoidális tér oldalán lévő membránokat réteg borítja lapos sejtek glia természet.
Az életkorral összefüggő változások
Változások a központi idegrendszerben in öreg kor elsősorban az agyi erek szklerotikus elváltozásaihoz kapcsolódik. Idős korban megvastagszik az agy pia materje és arachnoid membránja. Mészlerakódások jelenhetnek meg bennük. Kortikális atrófia figyelhető meg agyféltekék, elsősorban a frontális és a parietális lebeny. A kéreg egységnyi térfogatára jutó neuronok száma csökken, ez elsősorban a sejthaláltól függ. A neuronok mérete csökken, részben elveszítik bazofil anyagukat, a sejtmagok sűrűbbé válnak, körvonaluk egyenetlenné válik. A motoros kéreg V. rétegének piramisai és a kisagykéreg piriform sejtjei gyorsabban változnak, mint mások. A lipofuscin granulátumok az idegrendszer különböző részeinek neuronjaiban halmozódnak fel.
Az összes legfontosabb emberi viselkedési reakció a központi idegrendszer segítségével valósul meg. A központi idegrendszer fő funkciói a következők:
- a test minden részének egyesítése egységes egésszé és azok szabályozása;
- állapot- és viselkedésmenedzsment testület a feltételeknek megfelelően külső környezetés az ő igényeit.
Magasabbrendű állatokban és emberekben A központi idegrendszer vezető része az agykéreg . Az emberi élet legösszetettebb funkcióit irányítja mentális folyamatok(tudat, gondolkodás, beszéd, emlékezet stb.).
A központi idegrendszer funkcióinak vizsgálatának fő módszerei az eltávolítás és irritáció módszerei (klinikán és állatokon), az elektromos jelenségek rögzítése, a módszer feltételes reflexek.
A központi idegrendszer vizsgálatára továbbra is új módszereket fejlesztenek ki: az ún komputertomográfia morfofunkcionális változásokat láthatunk az agyban különböző mélységekben; az infravörös sugárzásban történő fényképezés (hőképalkotás) lehetővé teszi az agy „legforróbb” pontjainak észlelését; Új adatokat szolgáltat az agy működéséről a mágneses oszcillációinak vizsgálata.
A neuronok alapvető funkciói és kölcsönhatásai
Az idegrendszer fő szerkezeti elemei az idegsejtek vagy neuronok.
A neuronok alapvető funkciói
A neuronokon keresztül az információ az idegrendszer egyik részéből a másikba kerül, információcsere zajlik az idegrendszer és a test különböző részei között. A legösszetettebb információfeldolgozási folyamatok az idegsejtekben játszódnak le. Segítségükkel kialakulnak válaszokat test (reflexek) külső és belső ingerekre.
És így, a neuronok fő funkciói: külső ingerek észlelése receptor funkció, feldolgozásuk integratív funkció és az idegi hatások átvitele más idegsejtekre vagy különféle munkaszervekre effektor funkció. Az információfeldolgozás fő folyamatai az idegsejt, vagyis a szóma testében zajlanak. Számos faszerű elágazó hajtás dendritek(görögül dendron fa) neuron bemenetként szolgál, amelyen keresztül a jelek bejutnak az idegsejtbe. A neuron kimenete a sejttestből kiinduló folyamat axon(görög tengely tengely), amely az idegimpulzusokat továbbítja egy másik idegsejtnek vagy működő szervnek (izom, mirigy). Az axon kezdeti része és megnyúlása a sejttestből való kilépés helyén különösen erősen ingerelhető. axondomb idegsejt. A sejtnek ebben a szegmensében keletkezik az idegimpulzus.
A neuronok típusai
A neuronok három fő típusra oszthatók: afferens, efferens és intermedier. Afferens neuronok(érzékeny vagy centripetális) információt továbbítanak a receptoroktól az 1CNS felé. Ezeknek a neuronoknak a teste a központi idegrendszeren kívül található gerinccsomókés a csomópontoknál agyidegek. Az afferens neuronoknak egy hosszú folyamata van - egy dendrit, amely a periférián érintkezésben van egy perceptív képződménnyel - egy receptorral vagy maga alkot receptort, valamint egy második folyamat - egy axon, amely bejut. hátsó szarvak a gerincvelőbe.
Efferens neuronok(centrifugális) a leszálló hatások átvitelével járnak az idegrendszer fedőrétegeiből az alatta lévőkbe vagy a központi idegrendszerből a működő szervekbe. Az efferens neuronokat rövid folyamatok - dendritek - elágazó hálózata és egy hosszú folyamat - egy axon - jellemzi.
Köztes neuronok(interneuronok vagy interneuronok) ezek általában kisebb sejtek, amelyek különböző (különösen afferens és efferens) neuronok között kommunikálnak. Az idegi hatásokat vízszintes irányban (például a gerincvelő egyik szegmensén belül) és függőleges irányban (például a gerincvelő egyik szegmenséből a többi feletti vagy alatta lévő szegmensbe) továbbítják. Az axon számos ága miatt az interneuronok egyidejűleg sok más neuront is gerjeszthetnek.
Serkentő és gátló szinapszisok
A neuronok egymással (és az effektorszervekkel) való kölcsönhatása keresztül megy végbe speciális oktatás szinapszisok (görögül: kontaktus). Egy neuron testén lévő terminális ágai vagy egy másik neuron folyamatai alkotják őket. Minél több szinapszis van egy idegsejtben, annál jobban érzékeli a különféle irritációkat, és ezért annál szélesebb a befolyása a tevékenységére és a test különböző reakcióiban való részvétel lehetőségére. Különösen sok szinapszis van az idegrendszer magasabb részein és pontosan a legösszetettebb funkciójú neuronokban.
A szinapszis szerkezetében három elem található (1. ábra):
- preszinaptikus membrán, az axon terminális ágának membránjának megvastagodásával keletkezik;
- szinaptikus hasadék neuronok között;
- posztszinaptikus membrán a következő neuron szomszédos felületének megvastagodása.
Rizs. 1. Szinapszis diagram
Elő. preszinaptikus membrán,
Gyors. posztszinaptikus membrán,
C szinoptikus buborékok,
szinoptikus szakadék,
M mitokondrium,
Ó, acetilkolin
P receptorok és pórusok (pórusok)
dendrit (D) következő
idegsejt.
Nyíl egyoldalú gerjesztés.
A legtöbb esetben a befolyás átvitele egyik neuronról a másikra kémiai úton történik. A kontaktus preszinaptikus részében vannak szinaptikus vezikulák , amelyek speciális anyagokat tartalmaznak közvetítők vagy közvetítők. Ezek lehetnek acetilkolin (a gerincvelő egyes sejtjeiben, a vegetatív csomópontokban), noradrenalin (a szimpatikus idegrostok végződéseiben, a hipotalamuszban), néhány aminosavak stb. Az axonvégződésekre érkező idegimpulzusok a szinaptikus vezikulák kiürülését és a transzmitter szinaptikus hasadékba való kijutását okozzák.
A következő idegsejtre gyakorolt hatás természete alapján serkentő és gátló szinapszisokat különböztetnek meg.
Izgató szinapszisoknál A mediátorok (például az acetilkolin) a szinaptikus membrán specifikus makromolekuláihoz kötődnek, és depolarizációt okoznak. Ebben az esetben a membránpotenciál kismértékű és rövid távú (kb. 1 ms) fluktuációja a depolarizáció vagy serkentő posztszinaptikus potenciál(EPSP). Ahhoz, hogy a neuron gerjeszteni tudjon, az EPSP-nek el kell érnie egy küszöbszintet. Ehhez a membránpotenciál depolarizációs eltolódásának legalább 10 mV-nak kell lennie. A közvetítő hatása nagyon rövid ideig tart (1-2 ms), majd hatástalan komponensekre bomlik (pl. az acetilkolint a kolinészteráz enzim kolinná és ecetsav ) iszap, és a preszinaptikus terminálisok (például a noradrenalin) visszaszívják.
BAN BEN gátló szinapszisok tartalmazott gátló mediátorok (Például, gamma-amino-vajsav ). A posztszinaptikus membránra gyakorolt hatásuk a sejtből a káliumionok felszabadulását és a membrán polarizációjának fokozódását okozza. Ebben az esetben a membránpotenciál rövid távú ingadozása felé hiperpolarizáció gátló posztszinaptikus potenciál(TPSP). Ennek eredményeként az idegsejt gátolódik. Nehezebb felkelteni, mint eredeti állapotban. Ehhez erősebb stimulációra lesz szükség a depolarizáció kritikus szintjének eléréséhez.
A neuron impulzusválaszának megjelenése
A test membránján és az idegsejt dendritjein egyaránt vannak serkentő és gátló szinapszisok . Bizonyos időpontokban ezek egy része inaktív lehet, míg a másik rész aktív hatással van a membrán szomszédos területeire. A neuron membránpotenciáljának általános változása annak az eredménye összetett interakció a helyi EPSP-k és az összes aktivált szinapszis IPSP-jének (integrációja). Mind a serkentő, mind a gátló szinapszisok egyidejű hatására hatásuk algebrai összegzése (vagyis kölcsönös kivonása) következik be. Ebben az esetben a neuron gerjesztése csak akkor következik be a serkentő posztszinaptikus potenciálok összege lesz több, mint az összeg gátló . Ennek a többletnek egy bizonyos küszöbértéknek kell lennie (kb. 10 mV). Csak ebben az esetben jelenik meg a sejt akciós potenciálja. Meg kell jegyezni, hogy általában egy neuron ingerlékenysége a méretétől függ: minél kisebb a sejt, annál nagyobb az ingerlékenysége .
Az akciós potenciál megjelenésével a lebonyolítás folyamata ingerület az axon mentén és továbbítja azt a következő neuronnak vagy működő szervnek, azaz. a neuron effektor funkcióját végzik. Az idegimpulzus a neuronok közötti kommunikáció fő eszköze.
És így, az információ átvitele az idegrendszerben két mechanizmus segítségével történik: elektromos (EPSP; IPSP; akciós potenciál) és vegyi (közvetítők).
Az idegközpontok tevékenységének jellemzői
Az idegközpontok tulajdonságai nagymértékben összefüggenek a különböző idegsejteket összekötő szinapszisokon keresztül az idegimpulzusok vezetésének jellemzőivel.
Az idegközpontokon keresztül történő gerjesztés jellemzői
Idegközpont egy funkció végrehajtásához szükséges idegsejtek gyűjteményének nevezik. Ezek a központok megfelelő reflexreakciókkal reagálnak a hozzájuk kapcsolódó receptoroktól kapott külső stimulációra. Az idegközpontok sejtjei a rajtuk átáramló vérben lévő anyagok közvetlen irritációjára is reagálnak (humorális hatások). Egy teljes szervezetben szigorú koordináció van tevékenységeik összehangolása.
A gerjesztési hullám szinapszison keresztül egyik idegsejtről a másikra vezetése a legtöbb idegsejtben kémiailag történik. mediátor segítségével, a mediátor pedig csak a szinapszis preszinaptikus részében található, a posztszinaptikus membránban hiányzik. Ezért fontos jellemzője a gerjesztés vezetése szinoptikus kontaktusokon keresztül az egyoldalú vezetés idegi hatások, ami csak a preszinaptikus membrántól a posztszinaptikus membránig lehetséges, ellenkező irányba pedig lehetetlen. Ebben a tekintetben az idegimpulzusok áramlása a reflexívben bizonyos irányú az afferens neuronoktól az interkaláris neuronokig, majd az efferens motoros neuronokig vagy az autonóm neuronokig.
Nagyon fontos az idegrendszer tevékenységében van egy másik jellemzője a gerjesztés szinapszisokon keresztül történő levezetése lassú vezetés. Az idegimpulzus preszinaptikus membránhoz való közeledésének pillanatától a posztszinaptikus membránban potenciálok megjelenéséig tartó folyamatokra fordított időt ún. szinaptikus késleltetés. A legtöbb központi neuronban körülbelül 0,3 ms. Ezt követően több időre van szükség a serkentő posztszinaptikus potenciál (EPSP) és az akciós potenciál kialakulásához. Az idegimpulzus teljes átviteli folyamata (az egyik sejt akciós potenciáljától a következő sejt akciós potenciáljáig) egy szinapszison keresztül körülbelül 1,5 ms-ot vesz igénybe. Fáradtság, lehűlés és számos egyéb hatás hatására a szinaptikus késleltetés időtartama megnő. Ha bármilyen reakció részvételt igényel nagyszámú neuronok (sok száz, sőt ezer), akkor az idegközpontokon keresztül történő vezetési késleltetés összértéke tizedmásodperc, sőt egész másodperc is lehet.
A reflex tevékenység során teljes idő a külső irritáció alkalmazásának pillanatától a szervezet válaszának megjelenéséig az ún a reflex rejtett vagy látens ideje főként a szinapszisokon keresztüli vezetés időtartama határozza meg. Fontos a reflex látens idejének nagysága indikátor funkcionális állapot idegközpontok . Egy személy külső jelre adott egyszerű motoros reakciójának látens idejének mérését széles körben használják a gyakorlatban a központi idegrendszer funkcionális állapotának felmérésére (2. ábra).
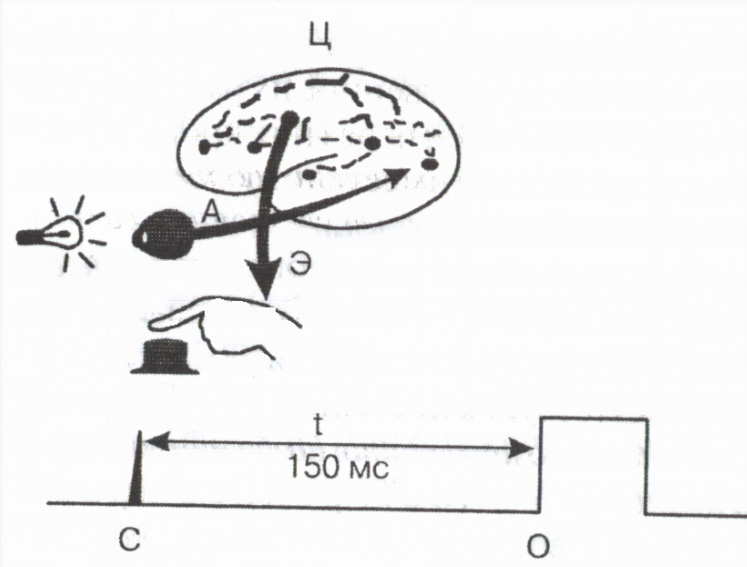
Rizs. 2. A motoros reakcióidő mérési sémája
Egy afferens
E efferens és C központi pályák; C - fényjelző jel,
O gombnyomás jel,
t 150 ms reakcióidő.
Gerjesztési összegzés
A receptoroktól az idegsejtekig terjedő egyetlen afferens hullámra válaszul kis mennyiségű transzmitter szabadul fel a szinapszis preszinaptikus részében. Ebben az esetben az EPSP általában a neuron posztszinaptikus membránjában fordul elő, kis helyi depolarizáció. Ahhoz, hogy a teljes EPSP érték a teljes neuronmembránon elérje az akciós potenciál előfordulásának küszöbét, sok küszöb alatti EPSP összegzésére van szükség a sejtmembránon. Csak a gerjesztés ilyen összegzésének eredményeként jön létre neuronválasz. Különbséget teszünk térbeli és időbeli összegzés között.
Térbeli összegzés több impulzus egyidejű vétele esetén figyelhető meg ugyanabba a neuronba különböző preszinaptikus rostok mentén. A szinapszisok egyidejű gerjesztése a neuronmembrán különböző részein a teljes EPSP amplitúdóját egy küszöbértékre növeli. Ennek eredményeként a neuronból válaszimpulzus keletkezik, és reflexreakció lép fel. Például egy gerincvelői motorsejt válaszának megszerzéséhez jellemzően 50-100 afferens rost egyidejű aktiválása szükséges a megfelelő perifériás receptoroktól.
Időösszegzés akkor fordul elő, ha ugyanazt az afferens útvonalat egymást követő ingerek sorozata aktiválja. Ha a beérkező impulzusok közötti intervallumok kellően rövidek, a korábbi ingerekből származó neuron EPSP-jének nincs ideje lecsengeni, akkor a következő EPSP-k egymásra helyeződnek, amíg a neuron membrán depolarizációja el nem éri a cselekvés bekövetkezéséhez szükséges kritikus szintet. lehetséges. Ilyen módon akár enyhe irritációk bizonyos idő elteltével a szervezet válaszreakcióit válthatnak ki (például tüsszögést és köhögést a légutak nyálkahártyájának enyhe irritációjára).
A ritmus átalakulása és asszimilációja
Egy neuron válaszkisülésének jellege nemcsak az inger tulajdonságaitól függ, hanem magának az idegsejtnek a funkcionális állapotától is.(membrántöltése, ingerlékenysége, labilitása). Az idegsejteknek megvan az a tulajdonságuk, hogy megváltoztatják az átvitt impulzusok frekvenciáját, azaz. a ritmus transzformáció tulajdonsága.
Ha egy neuron erősen ingerelhető (például koffein bevétele után), megnövekedett impulzusfrekvencia (ritmusanimáció) , és alacsony ingerlékenységgel (például fáradtsággal) fordul elő lassítja a ritmust , mivel több bejövő impulzust össze kell összegezni, hogy végre elérjük az akciós potenciál fellépésének küszöbét. Az impulzusok gyakoriságának ezen változásai erősíthetik vagy gyengíthetik a szervezet külső ingerekre adott válaszait.
Ritmikus stimulációval egy neuron aktivitása a bejövő impulzusok ritmusára tud hangolódni, azaz megfigyelhető ritmuselsajátítás jelensége(Ukhtomsky A. A., 1928). A ritmuselsajátítás fejlesztése biztosítja számos idegközpont tevékenységének hangolása összetett motoros aktusok kontrollálásakor ez különösen fontos a ciklikus gyakorlatok tempójának megőrzéséhez.
Folyamatok nyomon követése
Az inger befejezése után az idegsejt vagy idegközpont aktív állapota általában egy ideig folytatódik. A nyomkövetési folyamatok időtartama változó: rövid a gerincvelőben (néhány másodperc vagy perc), sokkal hosszabb az agy központjaiban (tíz percek, órák vagy akár napok), és nagyon hosszúak az agykéregben (akár több évtizedig) .
A neuronok zárt körein keresztül keringő impulzusok tiszta és rövid távú gerjesztési állapotot tudnak fenntartani az idegközpontban. A hosszan tartó rejtett nyomok sokkal összetettebbek. Feltételezhető, hogy az idegsejtben a nyomok hosszú távú megőrzése minden jellemző tulajdonságok Az ingerlés alapja a sejtet alkotó fehérjék szerkezetének megváltozása és a szinaptikus kapcsolatok átstrukturálása.
A rövid (akár 1 óráig tartó) pulzus-utóhatások az alapja az ún rövidtávú memória , és a sejtekben végbemenő szerkezeti és biokémiai átrendeződésekkel kapcsolatos hosszú távú nyomok képezik a kialakulásának alapját hosszú távú memória.
A központi idegrendszer tevékenységének koordinálása
A központi idegrendszer tevékenységének koordinációs folyamatai két idegi folyamat koordinációján alapulnak: a gerjesztés és a gátlás. A gátlás egy aktív idegi folyamat , amely megakadályozza vagy elnyomja az izgalmat.
A gátlási folyamat jelentősége a központi idegrendszerben
Az idegközpontok gátlásának jelenségét először I. M. Sechenov fedezte fel 1862-ben. Ennek a folyamatnak a jelentőségét az „Agy reflexei” (1863) című könyvében tárgyalta.
A békacombot savba mártva, és ezzel egyidejűleg az agy egyes részeit irritálva (pl. asztali só a diencephalon területére), I. M. Sechenov a gerincvelő „savas” reflexének éles késését, sőt teljes hiányát észlelte (a mancs visszahúzódása). Ebből arra a következtetésre jutott, hogy egyes idegközpontok jelentősen megváltoztathatják más központok reflexaktivitását, különösen a fedő idegközpontok gátolhatják az alacsonyabbak aktivitását. A leírt tapasztalat a név alatt vonult be az élettan történetébe Sechenov fékezés .
A gátló folyamatok az idegi tevékenység koordinációjának szükséges összetevői. Először, a gátlási folyamat korlátozza a gerjesztés terjedését a szomszédos idegközpontokon, ami hozzájárul a koncentrációjához az idegrendszer szükséges területein. Másodszor, egyes idegközpontokban más idegközpontok gerjesztésével párhuzamosan keletkező gátlási folyamat, ezáltal kikapcsolja a felesleges tevékenységeket Ebben a pillanatban szervek . Harmadszor, az idegközpontokban kialakuló gátlás megvédi őket a túlzott túlterheléstől a munkavégzés során, azaz védő szerepet tölt be.
Posztszinaptikus és preszinaptikus gátlás
Fékezési folyamat , ellentétben az izgalommal, nem terjedhet egy idegrost mentén ez mindig helyi folyamat a szinaptikus kapcsolatok területén. A származási hely alapján megkülönböztetünk preszinaptikus és posztszinaptikus gátlást.
Posztszinaptikus gátlás ezek gátló hatások, amelyek a posztszinaptikus membránban jelentkeznek. Leggyakrabban ez a fajta gátlás társul a központi idegrendszerben a speciális gátló neuronok. Ezek az interneuronok speciális típusai, amelyekben az axonterminálisok gátló transzmittert bocsátanak ki. Az egyik ilyen közvetítő az gamma-amino-vajsav (GAM K).
A gátló neuronokhoz közeledő idegimpulzusok ugyanazt a gerjesztési folyamatot váltják ki bennük, mint más idegsejtekben. Válaszul egy normális akciós potenciál terjed a gátló sejt axonja mentén. Más neuronokkal ellentétben azonban az axonvégződések nem serkentő, hanem gátló transzmittert bocsátanak ki. Ennek eredményeként a gátló sejtek gátolják azokat a neuronokat, amelyeken az axonjaik végződnek.
A speciális gátló neuronok közé tartoznak a Renshaw-sejtek a gerincvelőben, a Purkinje-sejtek a kisagyban, a kosársejtek a diencephalonban stb. Például a gátló sejtek nagy jelentőséggel bírnak az antagonista izmok aktivitásának szabályozásában: az antagonista izmok ellazulásához vezetnek, ezáltal elősegítik az agonista izmok egyidejű összehúzódását (3. ábra).
Renshaw sejtek részt vesz a gerincvelő egyes motoros neuronjainak aktivitási szintjének szabályozásában. Amikor egy motoros neuront gerjesztenek, impulzusok jutnak el az axonja mentén az izomrostokhoz, és ezzel egyidejűleg az axon kollaterálisai mentén a Renshaw-gátló sejthez. Ez utóbbi axonjai „visszatérnek” ugyanahhoz a neuronhoz, ennek gátlását okozva. Minél több serkentő impulzust küld egy motoros neuron a perifériára (és így a gátló sejtre), annál erősebb ez visszatérő fékezés(a posztszinaptikus gátlás egy fajtája). Ilyen zárt rendszer a neuron önszabályozásának mechanizmusaként működik, megvédve a túlzott aktivitástól.
A kisagy Purkinje sejtjei a kéreg alatti sejtmagok és szárstruktúrák sejtjeire gyakorolt gátló hatásukkal részt vesznek az izomtónus szabályozásában.
Kosársejtek a dicephalonban olyan kapuk, amelyek lehetővé teszik vagy nem engedik a test különböző területeiről az agykéregbe jutó impulzusokat.
Preszinaptikus gátlás szinaptikus érintkezés előtt fordul elő a preszinaptikus régióban. A gátló idegsejt axonjának vége a serkentő idegsejt axonjának végén szinapszist hoz létre, ami ennek az axonnak a membránjának túlzottan erős depolarizációját okozza, ami gátolja az itt áthaladó akciós potenciálokat, és ezáltal gátolja az idegsejtek átvitelét. gerjesztés. Ez a fajta gátlás korlátozza az afferens impulzusok áramlását az idegközpontokba, és kikapcsolja a fő tevékenységen kívüli hatásokat.
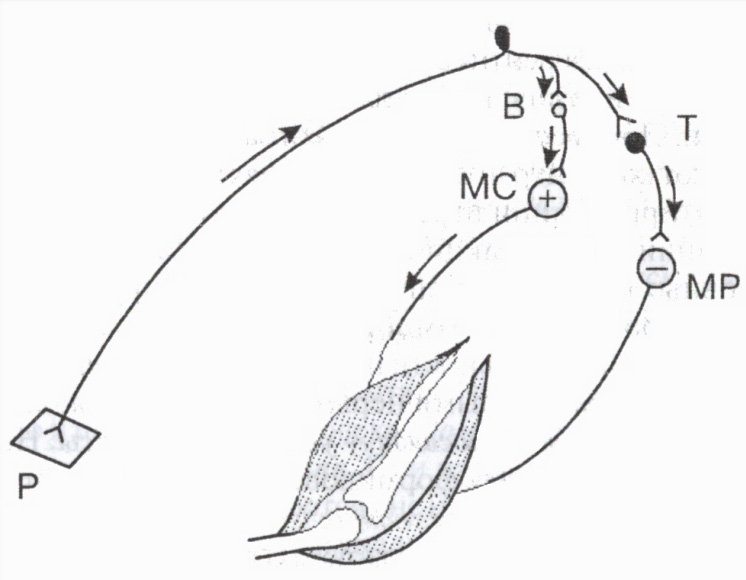
Rizs. 3. A gátló sejt részvétele az antagonista izmok szabályozásában
B és T serkentő és gátló neuronok. A hajlító izom (MS) motoros neuronjának gerjesztése (+) és az extensor izom motoros neuronjának gátlása (-) (MR). P bőrreceptor.
A besugárzás és a koncentráció jelensége
Ha egy receptort stimulálnak, a gerjesztés elvileg bármely irányba és bármely idegsejt felé terjedhet a központi idegrendszerben. Ez az egyik neuronjainak számos összekapcsolódása miatt következik be reflexív más reflexívek neuronjaival. A gerjesztési folyamat átterjedését más idegközpontokba jelenségnek nevezzük sugárzás.
Minél erősebb az afferens stimuláció és minél nagyobb a környező neuronok ingerlékenysége, annál több neuront fed le a besugárzási folyamat. A gátlási folyamatok korlátozzák a besugárzást, és hozzájárulnak a gerjesztés koncentrációjához a központi idegrendszer kiindulási pontjában.
A besugárzás folyamata fontos szerepet játszik pozitív szerepet a szervezet új reakcióinak kialakulása során (indikatív reakciók, kondicionált reflexek). Minél több különböző idegközpont aktiválódik, annál könnyebben kiválasztható közülük a későbbi tevékenységekhez leginkább szükséges központok. A besugárzásnak köszönhetően gerjesztés a különböző idegközpontok között új funkcionális kapcsolatok feltételes reflexek keletkeznek . Ezen az alapon lehetőség nyílik például új motoros készségek kialakítására.
Ugyanakkor a gerjesztés besugárzása is előfordulhat negatív hatás a test állapotára és viselkedésére, megzavarva az izgatott és gátolt idegközpontok közötti finom kapcsolatokat, és a mozgások koordinációjának zavara .
Uralkodó
Az intercentrális kapcsolatok sajátosságait vizsgálva A. A. Ukhtomsky felfedezte, hogy ha az állat testében komplex reflexreakciót hajtanak végre, például ismétlődő nyeléseket, akkor a motoros központok elektromos stimulációja nem csak ekkor szűnik meg a végtagok mozgását okozni. pillanatban, hanem fokozza a megindult láncreakció lefolyását, a nyelést, amely dominánsnak bizonyult.
A központi idegrendszerben a gerjesztés ilyen domináns fókuszát, amely meghatározza a test aktuális aktivitását, A. A. Ukhtomsky (1,923) jelölte meg a domináns kifejezéssel.
Domináns fókusz akkor fordulhat elő, ha emelt szint idegsejtek ingerlékenysége, amelyet különféle humorális és idegi hatások. Elnyomja más központok aktivitását, ezzel összefüggő gátlást fejt ki.
Nagyszámú neuron egy domináns rendszerré egyesülése az általános tevékenységi ütemre való kölcsönös ráhangolódás, azaz a ritmus asszimilációja révén történik. Egyes idegsejtek csökkentik magasabb aktivitási ütemüket, míg mások alacsony ütemüket egy bizonyos átlagos, optimális ritmusra növelik. A domináns sokáig rejtett, nyomkövető állapotban maradhat (potenciális domináns). Az előző állapot vagy a korábbi külső helyzet visszaállításakor újra felléphet a domináns (a domináns frissítése). Például a start előtti állapotban mindazok az idegközpontok aktiválódnak, amelyek a korábbi edzés során a munkarendszer részei voltak, és ennek megfelelően a munkával kapcsolatos funkciók felerősödnek. Szellemi kivégzés testmozgás vagy a mozgásábrázolás is reprodukálja a munkadominánst, amely a mozgásábrázolás oktató hatását biztosítja és az ún. ideomotoros tréning alapja. Teljes kikapcsolódással (pl autogén tréning) a sportolók törekednek a működő dominánsok megszüntetésére, ami felgyorsítja a felépülési folyamatokat.
Viselkedési tényezőként a domináns a legmagasabbhoz kapcsolódik ideges tevékenységés az emberi pszichológia. A domináns az élettani alapja a figyelem aktusa. Domináns jelenlétében a külső környezet számos hatása kívül marad figyelmünkön, de azokat, amelyek különösen érdekelnek, intenzívebben ragadjuk meg és elemezzük. Így a domináns erőteljes tényező a biológiailag és társadalmilag legjelentősebb ingerek kiválasztásában.
A gerincvelő és az agy kéreg alatti részeinek funkciói
A központi idegrendszerben több ősi szegmentális és evolúciósan fiatalabb szupraszegmentális az idegrendszer részei. A szegmentális szakaszok közé tartozik a gerincvelő, a medulla oblongata és a középagy, amelyek szakaszai szabályozzák az azonos szinten fekvő testrészek működését. Szuprasszegmentális szakaszok - a diencephalon, a cerebellum és az agykéreg nincs közvetlen kapcsolatban a test szerveivel, hanem az alatta lévő szegmentális szakaszokon keresztül irányítják tevékenységüket.
Gerincvelő
A gerincvelő a központi idegrendszer legalacsonyabb és legősibb része . Az emberi gerincvelő szürkeállománya körülbelül 13,5 millió idegsejtet tartalmaz. Tőlük a legtöbb (97%) köztes sejtek (interneuronok vagy interneuronok) , amelyek komplex koordinációs folyamatokat biztosítanak a gerincvelőn belül. A gerincvelő motoros neuronjai között vannak nagyok alfa motoros neuronokés kicsiket gamma motoros neuronok. A motoros idegek legvastagabb és leggyorsabban vezető rostjai az alfa motoros neuronoktól távoznak, összehúzódásokat okozva csontváz izomrostok. A gamma-motoros neuronok vékony rostjai nem okoznak izomösszehúzódást. Megközelítik a pro-prioreceptorokat izomorsókés szabályozzák érzékenységüket.
A gerincvelői reflexek részre osztható motor az elülső szarvak alfa motoros neuronjai végzik, és vegetatív az oldalsó szarvak afferens sejtjei végzik.
A gerincvelő motoros neuronjai az összes vázizmot beidegzik (kivéve az arcizmokat) . A gerincvelő végzi elemi motoros reflexek hajlítás és nyújtás, ritmikus, léptető, a bőr vagy az izmok és inak proprioceptorainak irritációjából eredő, valamint állandó impulzusokat küld az izmoknak, támogatva izomtónus. A speciális motoros neuronok beidegzik a légzőizmokat – a bordaközi izmokat és a rekeszizomzatot, légzési mozgások. Az autonóm idegsejtek beidegzik az összes belső szervet (szívet, ereket, verejtékmirigyek, mirigyek belső szekréció, emésztőrendszer, húgyúti rendszer).
Vezető funkció A gerincvelő a perifériáról érkező információáramlásnak az idegrendszer fedő részei felé történő átvitelével, valamint az agyból a gerincvelőbe érkező impulzusok továbbításával jár.
Mögött utóbbi évek fejlett speciális technikák a gerincvelő aktivitásának tanulmányozására ben egészséges ember. Például az alfa motoros neuronok funkcionális állapotát a perifériás stimuláció során bekövetkező izomválaszpotenciálok változásaival értékelik - az ún. H-reflex (Hoffmann reflex) lábikra izom amikor ingerült sípcsont idegés által T-reflex (ín ínből) a talpizom az Achilles-ín irritációjával. Módszereket dolgoztak ki a gerincvelőn keresztül az agyba jutó potenciálok rögzítésére (ép testfelületekről).
Medulla oblongata és híd
A medulla oblongata és a híd (együtt a hátsó agy) az agytörzs része. A koponyaidegek nagy csoportja (V-től XII párig) beidegzi a bőrt, a nyálkahártyákat, a fej izmait és számos idegrendszert. belső szervek(szív, tüdő, máj). Itt vannak számos emésztési reflex központja rágás, nyelés, a gyomor és a belek egy részének mozgása, emésztőnedvek elválasztása, valamint egyes védőreflexek központjai (tüsszögés, köhögés, pislogás, könnyezés, hányás) és a víz-só és cukor anyagcsere központjai . Az IV kamra alján a medulla oblongatában egy létfontosságú légzőközpont található, amely belégzési és kilégzési központokból áll. Kis sejtekből áll, amelyek impulzusokat küldenek a légzőizmoknak a gerincvelő motoros neuronjain keresztül.
Közelében található szív- és érrendszeri központ . Nagy sejtjei szabályozzák a szív működését és az erek lumenét. A légző- és kardiovaszkuláris központok sejtjeinek összefonódása biztosítja azok szoros kölcsönhatását.
A medulla oblongata fontos szerepet játszik a motoros aktusok végrehajtásában és a vázizomzat tónusának szabályozásában, a nyújtóizmok tónusának növelése . Különösen a megvalósításban vesz részt testtartási igazító reflexek (nyaki, labirintus). A felszálló felszállók áthaladnak a medulla oblongatán a hallási, vesztibuláris, proprioceptív és tapintási érzékenység útjai .
Középagy
A középső agy a quadrigeminusból, a substantia nigrából és a vörös magokból áll. A quadrigeminus elülső gumóiban vannak vizuális szubkortikális központok , és hátul auditív . A középagy részt vesz a szemmozgások szabályozása , végrehajtani pupillareflex (sötétben pupillatágulás, világosban szűkület).
A quadrigeminálisok számos reakciót hajtanak végre, amelyek az orientáló reflex összetevői . Hirtelen irritációra a fej és a szemek az inger felé fordulnak, állatoknál a fülek hegyeznek. Ez a reflex (I. P. Pavlov szerint a „Mi ez?” reflex) szükséges ahhoz, hogy felkészítse a szervezetet minden új behatásra adott időben történő reakcióra.
Fekete anyag a középagy a rágási és nyelési reflexekhez kapcsolódik, részt vesz izomtónus szabályozása (főleg amikor csinálod apró mozdulatok ujjak) és a barátságos motoros reakciók megszervezésében.
Piros mag a középagy motoros funkciókat lát el szabályozza a vázizomzat tónusát , ami a hajlító izmok fokozott tónusát okozza. Jelentős hatással van a vázizomzat tónusára, a középagy számos telepítési reflexek a testtartás megtartásához (rektifikáló testfelhelyezés a koronával, stb.).
Diencephalon
A diencephalon magában foglalja a thalamust (vizuális thalamus) és a hypothalamust (subthalamus).
Keresztül thalamus áthaladnak minden afferens úton (a szagló kivételével), amelyek a kéreg megfelelő észlelési területeibe (halló, látás stb.) kerülnek. A talamusz magjai a következőkre oszlanak specifikus és nem specifikus . A konkrétak közé tartozik kapcsoló (relé) magok és asszociatív . A test összes receptoráról érkező afferens hatások a talamusz kapcsolómagjain keresztül jutnak el. Az asszociatív magok impulzusokat kapnak az átkapcsolódó magoktól, és biztosítják kölcsönhatásukat. Ezeken a magokon kívül a talamusz nem specifikus magokat is tartalmaz, amelyek aktiváló és gátló hatással is rendelkeznek a kéreg kis területein.
Kiterjedt kapcsolatainak köszönhetően a talamusz létfontosságú szerepet játszik a szervezet működésében. A talamuszból a kéregbe érkező impulzusok megváltoztatják a kérgi neuronok állapotát és szabályozzák a kortikális aktivitás ritmusa . A talamusz közvetlen részvételével történik feltételes reflexek kialakítása és motoros készségek fejlesztése, érzelmek kialakulása egy személy, az arckifejezései. A thalamusnak különösen nagy szerepe van az érzések fellépésében fájdalomérzések . Tevékenysége összefügg bioritmusok szabályozása egy személy életében (napi, szezonális stb.).
hipotalamusz a legmagasabb szubkortikális szabályozási központ vegetatív funkciók , Államok ébrenlét és alvás . Itt találhatók a vegetatív központok, anyagcsere szabályozása és a testet, biztosítva állandó testhőmérséklet fenntartása (melegvérű állatoknál) és normál szinten vérnyomás , támogatja víz egyensúly szabályozó éhség és jóllakottság érzése . A hipotalamusz hátsó magjainak irritációja a szimpatikus hatások növekedését okozza, az elülsők pedig paraszimpatikus hatásokat.
A hipotalamusz és az agyalapi mirigy kapcsolata miatt ( hipotalamusz-hipofízis rendszer) az endokrin mirigyek tevékenysége szabályozott. A hipotalamusz által szabályozott autonóm és hormonális reakciók az emberi érzelmi és motoros reakciók összetevői.
Nem specifikus agyi rendszer
Nem specifikus rendszer az agytörzs középső részét foglalja el. Nem jár semmilyen specifikus érzékenység elemzésével vagy specifikus reflexreakciók végrehajtásával. Az impulzusok ebbe a rendszerbe oldalsó ágakon keresztül jutnak be minden meghatározott útvonalról, ami kiterjedt kölcsönhatást eredményez. Mert nem specifikus rendszer Jellemző a neuronok elhelyezkedése diffúz hálózat formájában, folyamataik bősége és változatossága. Ebből a szempontból kapta a nevet retikuláris képződés vagy retikuláris képződés.
Megkülönböztetni kétféle befolyás nem specifikus rendszer más idegközpontok munkájára aktiváló és gátló . E hatások mindkét típusa lehet növekvő (a fedőközpontokhoz) és csökkenő (a mögöttes központokhoz). -ért szolgálnak az agy funkcionális állapotának szabályozása, az ébrenléti szint és a vázizmok testtartási-tonikus és fázisos reakcióinak szabályozása.
Kisagy
A kisagy egy szupraszegmentális képződmény, amelynek nincs közvetlen kapcsolata a végrehajtó apparátussal. A kisagy egy páratlan képződményből áll - a vermisből és a páros féltekékből.
A kisagykéreg fő idegsejtjei számosak Purkinje sejtek. A kiterjedt kapcsolatoknak köszönhetően (minden sejtben akár 200 000 szinapszis is van) tartalmaznak sokféle szenzoros hatás integrálása , elsősorban proprioceptív, tapintható és vestibularis. A kisagykéregben a különböző perifériás receptorok reprezentációja van szomatotop szervezet (görög szomatosz test, toposz hely), azaz az emberi szervezetben való elhelyezkedésük sorrendjét tükrözi. Ezenkívül ez az elrendezési sorrend megfelel az agykéregben a testrészek ábrázolásának azonos elrendezési sorrendjének, ami megkönnyíti a kéreg és a kisagy közötti információcserét, és biztosítja közös tevékenységüket az emberi viselkedés szabályozásában. A kisagyi neuronok helyes geometriai szerveződése határozza meg annak fontossága az időszámlálásban és a ciklikus mozgások tempójának egyértelmű tartásában.
A kisagy fő funkciója a testtartási reakciók szabályozása és a motoros aktivitás koordinálása.(Orbeli L.A., 1926).
Által anatómiai jellemzők(a kisagykéreg kapcsolatai a magjaival) és funkcionális jelentősége, a kisagy fel van osztva három hosszanti zóna:
- a vermis belső vagy mediális kérge, melynek feladata a vázizomzat tónusának szabályozása, a testtartás és az egyensúly megőrzése;
- közbülső középső része agykérget a kisagy, amelynek feladata a testtartási reakciók mozgásokkal való összehangolása és a hibák kijavítása;
- a féltekék laterális vagy laterális kérge kisagy, amely együtt diencephalon az agykéreg pedig a gyors ballisztikus mozgások (dobások, ütések, ugrások stb.) programozásában vesz részt.
Alapi idegsejtek
A bazális ganglionok közé tartozik a striatum, amely a caudatus magból és a putamenből, valamint a sápadt magból áll. amygdala(kapcsolatos vegetatív központok limbikus rendszer) és a középagy substantia nigra.
Az afferens hatások a test receptoraiból a thalamuson keresztül és az agykéreg minden területéről érkeznek a bazális ganglionokhoz. Szinte kizárólag a striatumba jutnak be. A belőle származó hatások a sápadt magba, majd tovább az extrapiramidális rendszer szárközpontjaiba, valamint a thalamuson keresztül vissza a kéregbe irányulnak.
A bazális ganglionok részt vesznek a kondicionált reflexek kialakításában és a komplex megvalósításában feltétlen reflexek(védelmi, élelmiszer-előállító stb.). Biztosítják a fizikai munkavégzés során szükséges testhelyzetet, valamint az automatikus ritmikus mozgások (ősi automatizmusok) áramlását.
A fő motoros funkciót a nucleus pallidus látja el, tevékenységét a striatum szabályozza. Jelenleg a nucleus caudatus fontossága az összetett mentális folyamatok – a figyelem, a memória és a hibaészlelés – szabályozásában feltárult.
A test minden funkciója szomatikusra osztható, vagy állat(állatok) az észleléssel kapcsolatos külső információés izomtevékenység, valamint vegetatív (növényi) a belső szervek tevékenységével, a légzési, vérkeringési, emésztési, kiválasztási, anyagcsere-, növekedés- és szaporodási folyamatokkal.
Az autonóm idegrendszer funkcionális szerveződése
Vegetativ idegrendszer a gerincvelő és az agy efferens idegsejtjei, valamint a belső szerveket beidegző speciális csomópontok (ganglionok) sejtjei. A különböző testreceptorok stimulálása mind a szomatikus, mind az autonóm funkciókban változásokat okozhat, mivel ezeknek a reflexíveknek az afferens és centrális szakasza gyakori. Csak a különböző szakaszaikban különböznek egymástól. Jellemző tulajdonság Az autonóm reflexek reflexíveiben szereplő efferens pályák a két neuronból álló szerkezetük(az egyik neuron a központi idegrendszerben, a másik a ganglionokban vagy a beidegzett szervben található).
Vegetatív idegrendszer két részre osztható: szimpatikus és paraszimpatikus (4. ábra).
Efferens utak szimpatikus idegrendszer a mellkasban kezdődik és ágyéki régiók gerincvelő oldalsó szarvának neuronjaiból. A gerjesztés átvitele a prenodális szimpatikus rostokról a posztnodális rostokra egy mediátor részvételével történik acetilkolin , illetve a posztnodális rostoktól a beidegzett szervekig mediátor közreműködésével noradrenalin . Kivételt képeznek azok a rostok, amelyek beidegzik a verejtékmirigyeket és kitágítják a vázizmok ereit, ahol a gerjesztést acetilkolin segítségével továbbítják.
Efferens utak paraszimpatikus idegrendszer az agyban kezdődik néhány magból a középső és medulla oblongata, a gerincvelőben pedig a neuronoktól szakrális régió. A gerjesztés vezetése a paraszimpatikus pálya szinapszisaiban közvetítő közreműködésével történik acetilkolin . A második efferens neuron a beidegzett szervben vagy annak közelében található.
Az autonóm funkciók legmagasabb szabályozója a hipotalamusz , amely a retikuláris formációval és az agykéreg irányítása alatt álló limbikus rendszerrel együtt hat. Ezenkívül magukban a szervekben vagy azokban elhelyezkedő neuronok szimpatikus csomópontok, a központi idegrendszer részvétele nélkül is végrehajthatják saját reflexreakcióikat "perifériás reflexek" .
Rizs. 4. Autonóm idegrendszer
Bal oldali szál kilépési terület: paraszimpatikus (fekete)
és szimpatikus (árnyékolt) rendszerek.
A jobb oldalon az autonóm reflexek reflexíve efferens részének felépítése látható. A bal oldalon a középső, a medulla oblongata és a gerincvelő diagramja látható.
Az arab számok a mellkasi szakaszok, a római számok az ágyéki szakaszok számai.
A szimpatikus idegrendszer funkciói
A szimpatikus idegrendszer részvételével a szervezetben számos fontos reflex jön létre, amelyek célja annak aktív állapotának biztosítása, beleértve a motoros aktivitást is. Ide tartoznak a reflexek a hörgők kitágulása, fokozott pulzusszám és felerősödés , a szív és a tüdő ereinek kitágulása, miközben összehúzza a bőr és a szervek ereit hasi üreg (a vér újraelosztásának biztosítása ), a raktározott vér felszabadulása a májból és a lépből, a glikogén lebomlása glükózzá a májban ( szénhidrát energiaforrások mobilizálása ), az endokrin mirigyek és a verejtékmirigyek fokozott aktivitása. Szimpatikus idegrendszer csökkenti számos belső szerv aktivitását: a vesék érszűkülete következtében a vizeletképződés folyamatai csökkennek, a szervek szekréciós és motoros aktivitása gátolt gyomor-bél traktus; a vizeletürítést megakadályozzák, a falizom ellazul Hólyagés a záróizma összehúzódik.
A szervezet fokozott aktivitását szimpatikus pupillatágító reflex . A test motoros aktivitása szempontjából nagy jelentősége van a szimpatikus idegek trofikus hatása a vázizmokra , javítják anyagcseréjüket és funkcionális állapotukat, enyhítik a fáradtságot.
A szimpatikus idegrendszer nemcsak a szervezet működési szintjét növeli, hanem mozgósítja rejtett funkcionális tartalékait , aktiválja az agyi aktivitást, fokozza a védekező reakciókat ( immunreakciók, gát mechanizmusok stb.), hormonális reakciókat vált ki. A szimpatikus idegrendszer különösen fontos fejlesztés során stressz körülmények , a legtöbbben nehéz körülményekélettevékenység. L. A. Orbeli hangsúlyozta létfontosságú rokonszenves hatások a test alkalmazkodásához (adaptációjához) a kemény munkához, ahhoz különböző feltételek külső környezet. Ezt a funkciót ő nevezte el adaptív-trófikus.
A paraszimpatikus idegrendszer funkciói
Paraszimpatikus idegrendszer végrehajtani a hörgők szűkülete, a szívösszehúzódások lassulása és gyengülése: a szív ereinek szűkülése; az energiaforrások feltöltése (glikogén szintézise a májban és az emésztési folyamatok erősítése); a vizeletképző folyamatok erősítése a vesékben valamint a vizelési aktus biztosítása (a húgyhólyag izomzatának összehúzódása és a záróizom ellazulása) stb. A paraszimpatikus idegrendszer elsősorban kiváltó hatások : a pupilla, a hörgők összehúzódása, az aktivitás aktiválása emésztőmirigyek stb.
Az autonóm idegrendszer paraszimpatikus részlegének tevékenysége az áramra irányul a funkcionális állapot szabályozása, az állandóság fenntartása belső környezet homeosztázis . A paraszimpatikus osztály gondoskodik különböző élettani mutatók helyreállítása , élesen megváltozott miután intenzív izommunka, az elhasznált energiaforrások pótlása. Közvetítő paraszimpatikus rendszer Az acetilkolin, amely csökkenti az adrenerg receptorok érzékenységét az adrenalin és a noradrenalin hatására, bizonyos stresszoldó hatás .
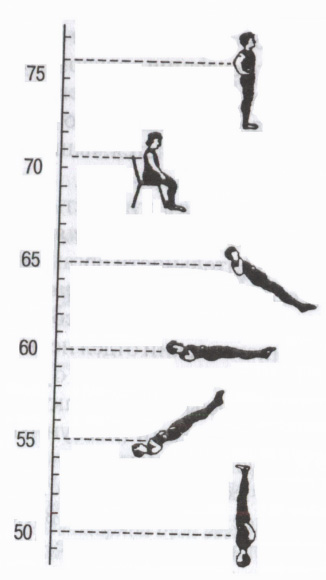
Rizs. 5. Autonóm reflexek
A testhelyzet hatása a pulzusszámra (bpm). (Mogendovich M.R. szerint, 1972)
Az autonóm szimpatikus és paraszimpatikus pályákon keresztül a központi idegrendszer bizonyos autonóm reflexeket hajt végre, a külső és belső környezet különböző receptoraitól kezdve: zsigeri-zsigeri (a belső szervektől a belső szervekig - például a légzés-szív reflex); dermo-zsigeri (a bőrből - a belső szervek aktivitásának változásai az irritáció során aktív pontok bőr, például akupunktúra, akupresszúra); a szemgolyó receptoraiból okulo-kardiális Aschner-reflex (szívverés csökkenése megnyomásakor szemgolyók paraszimpatikus hatás); motoros-zsigeri - pl. ortosztatikus teszt(fekvő helyzetből álló helyzetbe való mozgáskor megnövekedett pulzus szimpatikus hatás) stb. (5. ábra). A szervezet funkcionális állapotának és különösen az autonóm idegrendszer állapotának felmérésére szolgálnak (a szimpatikus vagy paraszimpatikus részleg befolyásának felmérésére).
Limbikus rendszer
Alatt limbikus rendszer megérteni számos kérgi és szubkortikális struktúrát, amelyek funkciói a szervezethez kapcsolódnak motivációs-érzelmi reakciók, memória és tanulási folyamatok .
A limbikus rendszer legmagasabb szakaszát képviselő kérgi szakaszok az agyféltekék alsó és belső felületein helyezkednek el (a frontális kéreg részei, gyrus vagy limbikus kéreg, hippocampus stb.). NAK NEK szubkortikális struktúrák A limbikus rendszer magában foglalja a hipotalamusz, a talamusz néhány magja, a középagy és a retikuláris képződés. Mindezen részlegek között szoros közvetlen és visszacsatolási kapcsolatok vannak, amelyek az úgynevezett „limbikus gyűrűt” alkotják.
A limbikus rendszer a szervezet tevékenységének legkülönfélébb megnyilvánulásaiban vesz részt: az étkezési-ivási viselkedés szabályozásában, az alvás-ébrenlét ciklusban, az emléknyom kialakulásának folyamataiban (lerakódás és az emlékezetből való előhívás), a fejlődésben. az agresszív-defenzív reakciók , biztosítva a szelektív viselkedést. Ő pozitív és negatív érzelmek minden motoros, vegetatív és hormonális komponenssel. A limbikus rendszer különböző részeinek elektromos stimulációja beültetett elektródákkal (állatkísérletek során, a klinikán betegek kezelésekor) feltárta a jelenlétet örömközpontok, pozitív érzelmek kialakítása, és nemtetszését, negatív érzelmeket formálva. Az emberi agy mélyszerkezeteinek ilyen pontjainak elszigetelt irritációja az „oktalan öröm”, „értelmetlen melankólia” és „megmagyarázhatatlan félelem” érzését váltotta ki.
Az agykéreg funkciói
Magasabbrendű emlősökben és emberekben a központi idegrendszer vezető része az agykéreg.
Kortikális neuronok
A kéreg egy 2-3 mm vastag szürkeállomány réteg, amely átlagosan körülbelül 14 milliárd idegsejtet tartalmaz. Jellemző rá az interneuron kapcsolatok bősége, melyek növekedése 18 éves korig, sőt esetenként tovább is tart.
A kérgi sejtek fő típusai a piramis és a csillag alakú neuronok. Csillagszerű neuronok az irritációk észlelésének folyamataihoz és a különböző piramis neuronok tevékenységeinek egyesítéséhez kapcsolódik.
Piramis neuronok ellátja a kéreg efferens funkcióját (főleg a piramispályán keresztül) és a viutricorticalis interakciós folyamatokat egymástól távol eső idegsejtek között. A legnagyobb piramissejtek, a Betz óriási piramisai az elülső központi gyrusban (a kéreg motoros területén) találhatók.
A kéreg funkcionális egysége az függőleges oszlop egymással összefüggő neuronok. A függőlegesen megnyúlt nagy piramissejtek a felettük és alattuk elhelyezkedő neuronokkal neuronok funkcionális társulásait alkotják. Súly A függőleges oszlop neuronjai ugyanazon afferens stimulációra (azonos receptorból) ugyanazzal a reakcióval reagálnak, és együttesen alkotják a piramis neuronok efferens válaszait. Igény szerint a függőleges oszlopok nagyobb formációkká kombinálhatók, lehetővé téve az összetett reakciókat.
Különféle kérgi mezők funkcionális jelentősége
Az egyes kérgi területek szerkezeti sajátosságai és funkcionális jelentősége szerint a teljes kéreg három fő csoportra oszlik: elsődleges, másodlagos és harmadlagos mezőkre (6. ábra).
Elsődleges mezők az érzékszervekhez és a periférián lévő mozgásszervekhez kapcsolódik. Ők biztosítják érzések megjelenése . Ide tartozik például a fájdalom és az izom-ízületi érzékenység mezője a kéreg hátsó központi gyrusában, a látómező az occipitalis régióban, a hallásmező a temporális régióban és a motoros mező az elülső központi gyrusban. Az elsődleges területeken magasan specializálódott determináns sejtek, ill detektorok, szelektíven csak bizonyos ingerekre reagál. Például a látókéregben vannak detektor-neuronok, amelyek csak a fény be- vagy kikapcsolásakor gerjesztődnek, csak egy bizonyos intenzitásra, meghatározott fényintervallumokra, bizonyos hullámhosszra stb. A kéreg elsődleges mezőinek megsemmisülésekor úgynevezett kérgi vakság, kortikális süketség stb.
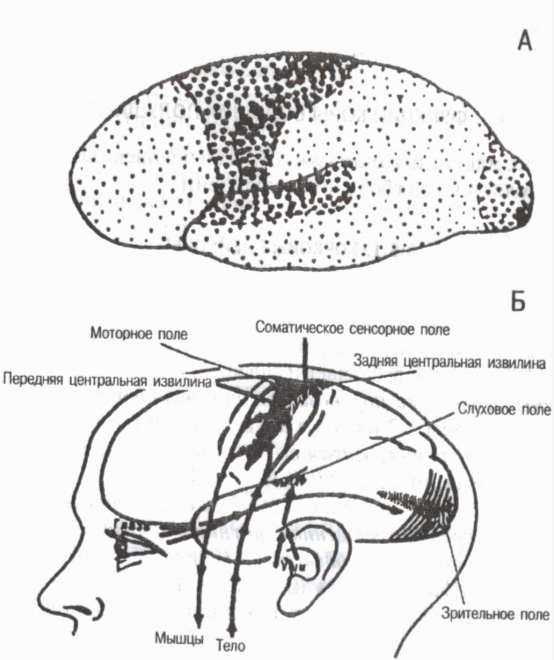
Rizs. 6. Az agykéreg elsődleges, másodlagos és harmadlagos mezői
A-n: nagypontos elsődleges mezők, közepes másodlagos mezők, kis tercier mezők (G. I. Polyakov 1964, A. R. Luria, 1971 szerint)
B-n: az agykéreg elsődleges (projekciós) mezői (V. Penfield, L. Robert szerint, 1964)
Másodlagos mezők az elsődlegesek mellett találhatók. Mi történik bennük a hang-, fény- és egyéb jelek megértése és felismerése, az általánosított észlelés összetett formái keletkeznek . Ha a másodlagos mezők megsérülnek, a tárgyak látásának és a hangok hallásának képessége megmarad, de a személy nem ismeri fel őket, és nem emlékszik a jelentésére.
Harmadlagos mezők szinte kizárólag emberben fejlődött ki. Ezek a kéreg asszociatív területei, amelyek az elemzés és szintézis magasabb formáit biztosítják, és céltudatos emberi viselkedési tevékenységet alakítanak ki. A harmadlagos mezők a cortex interparietalis, occipitalis és temporalis régióinak hátsó felében, az elülső részekben pedig az elülső felében helyezkednek el. frontális területek. Az ő szerepük különösen nagy mindkét félteke összehangolt munkájának megszervezésében . A harmadlagos mezők az emberben később érnek, mint más kérgi mezők, és az öregedés során korábban lebomlanak, mint mások.
A hátsó tercier mezők (főleg a kéreg inferior parietális területei) feladata az információ fogadása, feldolgozása és tárolása. Képet alkotnak arról test diagram és térdiagram , a mozgások térbeli orientációját biztosítva. Elülső tercier mezők (frontális területek) az emberi viselkedés összetett formáinak általános szabályozása, szándékok és tervek, önkéntes mozgalmak programjainak kialakítása, megvalósításuk nyomon követése . A harmadlagos mezők kialakulása az emberekben összefügg beszédfunkcióval. gondolkodni ( belső beszéd) csak különböző szenzoros rendszerek együttes tevékenységével lehetséges, amelyekből származó információk integrálása harmadlagos területeken történik. A harmadlagos mezők veleszületett fejletlensége esetén az ember nem képes elsajátítani a beszédet (csak értelmetlen hangokat ejt ki), és még a legegyszerűbb motoros készségeket sem (nem tud öltözni, szerszámokat használni stb.).
Páros aktivitás és féltekei dominancia
Ennek eredményeként információfeldolgozás történik mindkét félteke páros tevékenysége agy. Azonban, mint szabály, az egyik félteke vezető domináns . A legtöbb ember vezetésével jobb kéz(jobbkezes) A domináns a bal félteke, az alárendelt (szubdomináns) pedig a jobb félteke.
Bal félteke jobbhoz képest finomabb idegi szerkezettel, gazdagabb neuronkapcsolatokkal, koncentráltabb funkciókat és Jobb körülmények vérellátás A bal domináns féltekén található egy motoros beszédközpont (Broca központja), amely biztosítja beszédtevékenységés a beszéd érzékszervi központja, amely megérti a szavakat. A bal félteke a kézmozgások finom szenzomotoros szabályozására specializálódott.
Az emberben három formája van funkcionális aszimmetria: motoros, érzékszervi és mentális . Általában az embernek van egy domináns karja, lába, szeme és füle. A funkcionális aszimmetria problémája azonban meglehetősen összetett. Például egy jobbkezesnek lehet domináns bal szeme ill bal fül, amelyekből származó jelek dominánsak. Sőt, minden féltekén nemcsak az ellenkező, hanem a test ugyanazon oldalának funkciói is reprezentálhatók. Ennek eredményeként lehetséges az egyik félteke cseréje a másikkal sérülés esetén, és létrehoz szerkezeti alapja a változó féltekei dominanciának a motoros vezérlésben.
A mentális aszimmetria egy bizonyos formájában nyilvánul meg féltekei szakirányok. Mert bal agyfélteke elemző folyamatok, információk szekvenciális feldolgozása, beleértve a beszéd segítségével, az absztrakt gondolkodás, az átmeneti kapcsolatok értékelése, a jövőbeli események előrejelzése, a verbális és logikai problémák sikeres megoldása. BAN BEN jobb agyfélteke az információkat holisztikusan, szintetikusan (részletekre bontás nélkül), a múltbeli tapasztalatokat figyelembe véve, a beszéd részvétele nélkül dolgozzák fel, az érdemi gondolkodás dominál. Ezek a tulajdonságok lehetővé teszik a térbeli sajátosságok észlelését és a térlátási problémák megoldását a jobb agyféltekéhez társítani. A jobb agyfélteke funkciói a múlthoz, a bal félteke a jövőhöz kapcsolódnak.
Az agykéreg elektromos aktivitása
A kéreg funkcionális állapotában bekövetkezett változások tükröződnek a rögzítésében elektromos tevékenység elektroencefalogram (EEG). A modern elektroencefalográf 2-3 milliószorosára erősíti fel az agypotenciálokat, és lehetővé teszi az EEG egyidejű vizsgálatát a kéreg számos pontjáról, azaz tanulmányozását. rendszerfolyamatok. EEG regisztráció tintával papírra írt formában, valamint formában teljes kép az agy felszínének diagramján, azaz agytérképeken (térképezési módszer) a modern számítógépes rendszerek monitor képernyőjén (7. ábra).
Vannak bizonyos frekvencia tartományok , hívott EEG ritmusok (8. ábra): relatív nyugalmi állapotban leggyakrabban az alfa ritmus rögzíthető (8-13 oszcilláció 1 s-onként); aktív figyelem állapotában - béta ritmus (14 oszcilláció 1 másodpercnél és magasabbnál); elalváskor néhány érzelmi állapotok béta ritmus (4-7 oszcilláció 1s-onként); mélyalvásban eszméletvesztés, érzéstelenítés, delta ritmus (1 s-onként 1-3 fluktuáció).
Az EEG a kérgi neuronok interakciójának sajátosságait tükrözi a mentális és fizikai munka (Livanov M.N., 1972). A jól bevált koordináció hiánya szokatlan vagy nehéz munkavégzés során az ún EEG deszinkronizálás gyors aszinkron tevékenység. A motoros készség fejlődésével jelenségek jelennek meg az EEG-ben EEG szinkronizálás a kéreg különböző mozgásszabályozásában részt vevő területei elektromos aktivitásának összekapcsoltságának (szinkron és in-phase) növelése. Ciklikus munkavégzés során lassú potenciálok jelennek meg az elvégzett, képzeletbeli vagy közelgő mozgás ütemében "kijelölt ritmusok"(Sologub E.B., 1973).
Az EEG háttértevékenysége mellett külön az eseményekhez kapcsolódó lehetőségek: kiváltott potenciálokat, külső ingerekre válaszul (hallási, vizuális stb.); Az agyi folyamatokat tükröző potenciálok az egyes motoros folyamatok előkészítése, megvalósítása és befejezése során hat erre "várakozás hulláma" vagy kondicionált negatív hullám (Walter G., 1966), előmotoros, motoros és végső potenciálok stb. Ezen kívül különféle ultralassú oszcillációk néhány másodperctől több tíz percig tartó (különösen az úgynevezett „omega potenciálok” stb.), amelyek a funkciók és a mentális aktivitás szabályozásának biokémiai folyamatait tükrözik.

Rizs. 7. Agytérképezés
Humán elektroencefalogram (EEG) többcsatornás rögzítése a monitor képernyőjén és a kéreg gerjesztett (világos zónák) és gátolt (sötét zónák) területeinek visszaverődése.
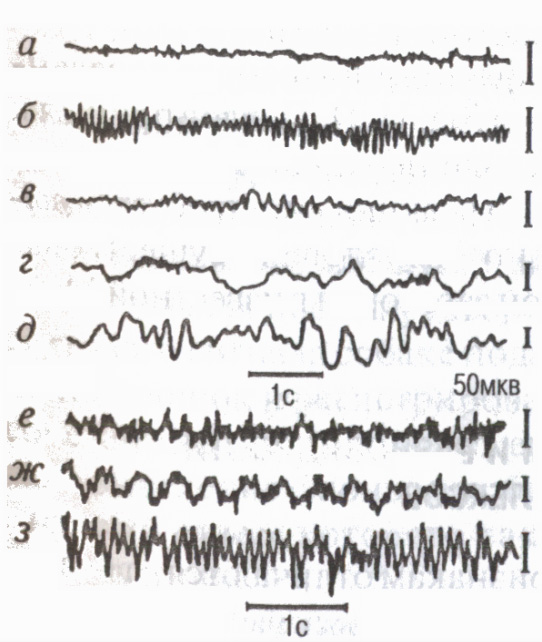
Rizs. 8. Az emberi agykéreg occipitalis (a-e) és motoros (f-h) területének EEG vizsgálata során különféle államokés izommunka során
aktív állapot,
nyitott szemek (béta ritmus); b- pihenés, csukott szemek (alfa ritmus);
c álmosság (tesha-ritmus);
d elalvás; d mély álom"H
(delta ritmus); e szokatlan ill
kemény munka aszinkron gyakori
aktivitás (a deszinkronizáció jelensége);
g ciklikus munka lassú potenciálok a mozgások ütemében ("címkézett ritmusok" EEG); h alfaritmus elsajátított mozgásmegjelenésének kivitelezése.
Gyógyászat és állatgyógyászat
Piramis neuronok. Nagy sejtek, perikarya 10-130 mikron méretű. A sejtben van egy apikális dendrit, amely a molekuláris réteg felé irányul; oldalirányú folyamatok – dendritek; egy hosszú axon származik az alapból - ez egy motoros idegrost lesz (a piramispálya kialakulásának kezdete).
Szövettan előadás 13. sz
Központi idegrendszer. Agykérget. Az agykéreg rétegeinek citoarchitektúrája. Neuron összetétele. A piramis neuronok jellemzői. Az agykéreg modul szerkezeti és funkcionális egysége. Kisagy. A kisagykéreg szerkezete és idegi összetétele.
Az agy szürke- és fehérállományának fejlődési forrása
Az idegcső szürkeállományának köpenyéből vagy köpenyzónájából fejlődik ki
Peremfátyol fehérállomány
Alkatrészek az agy szürkeállománya (neocortex)
6 réteg:
- A molekulárist főként az alatta lévő rétegek alatti sejtfolyamatok alakítják ki, de vannak olyan neuronok is, amelyeket vízszintesnek neveznek.
- Külső szemcsés jelenlét csillag alakú neuronok és nem sok piramis neuron.
- Külső piramis (piramis) a legszélesebb réteg, amelyet tartalmaz piramis neuronok.
- Belső szemcsés finom csillag alakú neuronok végeznek intrakortikális kapcsolatokat (nem lépnek be a fehérállományba).
- Belső piramis (ganglionális) képviseli nagy piramis alakú neuronok vagy Betts sejtek.
- A polimorf sejtformák változatosak, a sejtek kicsik, axonjaik a fehérállományba nyúlnak be.
Csillagszerű neuronok.A Perikarya kerek vagy háromszög alakú, átmérője 4-8 mikron; ezen sejtek axonja és dendritjei részt vesznek az intrakortikális kapcsolatokban.
Piramis neuronok.Nagy sejtek, 10-130 mikron méretű perikarya. A sejtben van egy apikális dendrit, amely a molekuláris réteg felé irányul; laterális folyamatok dendritek; egy hosszú axon az alapból származik, és egy motoros idegrost lesz (a piramispálya kialakulásának kezdete). A kollaterálisok az axonból származnak: a szomszédos sejtekhez mennek, vagy felemelkednek, és kölcsönhatásba lépnek a fedőrétegekkel.
Általános jellemzők kérgi neuronok:
Az agykéreg minden neuronja többpólusú. Funkció szerint 1-4 rétegek: asszociatív (interkaláris), 5-6 motoros.
Az agy szerkezeti és funkcionális egysége: modul
Modul (agykéregben 3 millióig) 300 mikron átmérőjű, a kéreg felületére merőleges függőleges oszlop, amely a kéreg teljes vastagságán áthalad.
A modul közepén:
- Az egyik corticocarticularis idegrost egy idegrost a kéregben; vagy egyazon félteke piramis neuronjaiból (asszociatív-interkaláris), vagy az ellenkező féltekéből (commisszurális) származik.
- Két thalamocorticalis rost (affirent-érzékeny rostok), amelyek a thalamus opticusból származnak és 4 rétegben futnak; akkor efferens idegrost keletkezik axonok alkotják A legjobb sejtek.
Citoarchitektúraa kéreg területei, amelyek a sejtek felépítésében és elhelyezkedésében különböznek egymástól.
Mieloarchitektúraa kéreg területei, amelyek a rostok elrendeződésének jellemzőiben különböznek egymástól.
Mezők a kéreg különböző területei, amelyek különböznek egymástól: cito, mieloarchitektúra és funkcionális jelentősége.
A kéreg fajtái:
- Szemcsés érzékeny központ, amelyben a 2. és 4. réteg jól fejlett; megszüntetik a szaglás-, hallás- és látásszervekből érkező (afferens) érzőidegrostokat.
- Agranuláris motoros központ, amelyben a 3., 5. és 6. réteg alakul ki.
Az intelligencia megbízható morfológiai megfelelője az asszociatív neuronok szinapszisainak száma (1-4 réteg), normál esetben 10 ezer.
Kisagy
A kisagy hátsó agyának fejlődési forrása
Funkciók: koordinálja a mozgást és az egyensúlyt
A kisagy nagyszámú fordulattal rendelkezik, elágazó fa formájában.
A gyri-ket barázdák választják el egymástól. Minden gyrus tartalmaz: egy keskeny fehér anyaglemezt, teljesen lefedve szürkeállomány(ugat). A neuroncsoportok mélyen a kisagy fehér anyagában helyezkednek el.: kisagyi magok.
A szürkeállomány vagy a kisagykéreg három rétegből áll:
- A külső molekulárist multipoláris asszociatív (interkaláris) neuronok képviselik. 2 típusú sejt létezik:
- A kosárneuronok a molekuláris réteg alsó harmadában, a sejtekben találhatók szabálytalan alakú kis méretek. A sejt axonja szinapszisban van a Purkenje sejt perikareonjával, ahol elágazik és kosár formájú plexust képez. Dendritek a molekuláris rétegben.
- A csillagképek (nagy és kicsi) a molekuláris réteg (a kéreg felszíne) külső részén lokalizálódnak.Nagy csillagneuronok:az axon szinapszisok a Purkenje sejtek perikarikájával és részt vesznek a kosár kialakításában. Dendritek a molekuláris rétegben.Kis csillagneuronok: Az axon szinapszisok a Purkenje sejtek dendritjeivel.
- Középső ganglion - egy réteg, a multipoláris motoros neuronok vagy Purkinje-sejtek testei képviselik. Ezek nagy körte alakú sejtek a perikarionból (test), amelyekben 2-3 dendrit nyúlik be a molekuláris rétegbe. Az axon a sejttestből származik, és a szemcsés rétegen keresztül a fehérállományba jut, ahol a kisagymagoknál végződik. A Purkinje sejt axonjai az egyetlen efferens (motoros) idegrostok, amelyek a kisagykéregből jönnek ki. Közelebb a sejttesthez az axonból kollaterálisok távoznak, amelyek a ganglionrétegbe és a molekuláris réteg mélyebb részeibe kerülnek, ahol szinapszisba lépnek a testekkel és a dendritekkel.szomszédos Purkinje sejteket.
- Belső szemcsés minden sejt poláris és asszociatív kétféle sejtet tartalmaz:
1-Sejt-szemcsék kis sejtek, perikaryonok 4-6 mikron átmérőjűek, dendritek: rövidek, 3-4 darab, a fehérállomány felé irányulnak, madárlábra emlékeztetnek. Az axon felemelkedik a molekuláris rétegbe, ahol T-alakban osztódik.
2-Nagy csillagkép: hosszú axonokkal és rövid axonokkal Második típusú Golgi-sejtek. A test nagy sejtjei közvetlenül a ganglionréteg alatt helyezkednek el. Az axon szinapszisok mohás rostjaival, mielőtt a szemcsesejtek dendritjeivel szinapszisba lépnének. A dendritek a molekuláris rétegbe emelkednek.
Afferens (érzékeny) idegrostok:
- Mohos szálak mohás terminál idegrost aljzatnak nevezik. A mohás rostrozetták a szemcsesejtek dendritjeivel szinapszisba lépnek, és kisagyi glomerulusokat alkotnak.
- Mászó vagy liána alakú rostok a fehérállományból származnak, áthaladnak a szemcsés rétegeken, és vagy a Purkinje sejt perikarionjával vagy e sejtek dendriteivel szinapszisba lépnek.
A szemcsesejtek axonjai dendritekkel szinapszisban lépnek fel:
- Sejt molekuláris réteg
- Purkenje sejtek
- Golgi sejt II
A kisagy gátló rendszere a következőket tartalmazza:
- A molekuláris réteg sejtjei
- 2-es típusú Golgi-sejtek
- Izgató szinapszisok:
- Mohos rostok
- Liana rostok
Valamint más művek, amelyek érdekelhetik |
|||
| 73026. | Adatok bevitele, formázása és képletek írása | 126 KB | |
| Cél: A Cellák formázása paranccsal formázza a D oszlopban lévő adatokat pénznemformátumban tizedesjegyek nélkül. A Formátum minta gombbal másolja a D oszlop formátumát E-be. A Formázás panel gombjaival adjon meg százalékos formátumot két tizedesjegygel az F oszlopban. | |||
| 73027. | Diagramok készítése és szerkesztése Excelben | 160 KB | |
| Tanuljon meg diagramokat készíteni a varázsló segítségével; Tanulj meg diagramokat szerkeszteni. Tesztkérdések Mi a diagramok célja Milyen típusú diagramokat ismersz Hogyan építsünk rá diagramot külön lap A diagram típusának megváltoztatása Diagram törlése... | |||
| 73028. | Fájlrendszerek modellezése | 147,5 KB | |
| A felhasználók szimbolikus neveket adnak a fájloknak, miközben figyelembe vesznek bizonyos operációs rendszer korlátozásokat. A könyvtár tartalmazza a benne található fájlok listáját, és megfeleltetést hoz létre a fájlok, valamint azok jellemzői és attribútumai között. | |||
| 73029. | A VNT-t helyettesítő NVM szerkezeti fázisösszetételének meghatározása röntgendiffrakciós és elektronmikroszkópos módszerekkel | 1,5 MB | |
| Ami fontos, az az NVM szerkezeti fázisú felépítése, hogy a VNT megfeleljen a röntgendiffrakciós és elektronmikroszkópos adatoknak. Figyelje meg az NVM szerkezeti fázisösszetételének változását a termokémiai feldolgozás során. | |||
| 73033. | Egyéni adattípusok példaként az MS SQL Server használatával | 51 KB | |
| Fedezze fel az egyéni adattípusok, DML és DDL triggerek, valamint integritási megszorítások meghatározásának és használatának lehetőségeit egy posztrelációs DBMS-ben. Mester módszerek és technológiák egyéni adattípusok létrehozására a CLR.Neten. | |||
| 73034. | A polgári épületek vasbeton födémeinek jelölésének elve | 47,1 KB | |
| A polgári épületek vasbeton födémeinek jelölésének elve A GOST 2300978 szabvány szerint a födémeket számos alfanumerikus index jelöli. Az első csoport tartalmazza a födém típusának megjelölését és tervezési méreteit, hosszát és szélességét deciméterben. | |||
Idegrendszer. 2. előadás.
Ugat A nagyagy (cortex cerebri) az agyféltekék felszínén összefüggő, 2-5 mm vastag szürkeállományréteg. Nemcsak a kanyarulatokon helyezkedik el, hanem minden barázdába benyúlik, aminek következtében területe viszonylag nagy, felnőttnél 2200 cm 2. A kéreg tömege 581 g, térfogata körülbelül 560 cm 3. Ebben az esetben a kéreg fő térfogata fehér anyagból áll (450 cm 3, súlya - 470 g). A kortikális neuronok sejttesttömege mindössze 21 g (20 cm3).
A kérgi idegsejtek mérete széles skálán mozog 8-9 között mk 150-ig mk. Az emberi agykéregben legfeljebb 15 milliárd idegsejt található, ebből 6 milliárd kis sejt. A kérgi neuronok túlnyomó többsége két típusba tartozik: a piramis neuronok és a stellate neuronok. Ezek a sejtek a kéregben találhatók bizonyos rétegekben.
Az emberi agyban számos filogenetikailag megkülönböztethető különböző típusok ugat:
Az ősi kéregnek - paleocortexnek (0,6%) - gyakorlatilag nincs rétegződése (1-2 réteg), a szaglóháromszögek területén lokalizálódik, a septum pellucidum borítását képezi, és körülveszi az amygdala magot.
A régi kéreg - archicortex (2,2%) - 2-3 rétegből áll, amelyet a gyrus fogfogának szürkeállománya, a hippocampus szürkeállománya és a corpus callosumon lévő medulláris striák képviselnek.
Átmeneti kéreg - mesocortex (1,6%) - a boltozatos gyrus területe.
Az új kéreg - neocortex (95,6%) - jól strukturált és 6 rétegű.
A magasabb gerincesekben túlnyomórészt hat többé-kevésbé egyértelműen meghatározott réteg található. De ezen rétegek mindegyike, az első kivételével, két vagy akár három alrétegre osztható.
Első réteg, az úgynevezett övezeti ill molekuláris lemez, főleg piramis neuronok csúcsi dendritjeinek plexusaiból áll, melyek sejttestei a kéreg más rétegeiben helyezkednek el. Az első rétegben nagyon kevés idegsejt található. Vízszintes sejteket tartalmaz, amelyek axonjai és dendritjei szintén vízszintesen ugyanabban a rétegben helyezkednek el.
Második réteg, az úgynevezett külső szemcsés ill külső szemcsés lemez, kis piramisokhoz, úgynevezett interneuronokhoz és csillagneuronokhoz tartozó kis sejtek tömegét foglalja magában, a második rétegben pedig a kis piramissejtek dominálnak.
A harmadik réteg a külső piramislemez közepes méretű piramissejteket tartalmaz
Negyedik réteg, az úgynevezett belső szemcsés ill belső szemcsés lemez, főleg kis csillagsejtekből áll, de kis és közepes méretű piramisokat is tartalmaz.
Az ötödik réteg a belső piramislemezóriás piramissejteket vagy Betz sejteket tartalmaz.
Hatodik réteg – többformájú lemez főként közepes méretű piramissejteket és kis számban kis piramis- és csillagsejteket tartalmaznak.
Az első három réteg a legfiatalabb, ezek biztosítják a kapcsolatot a kéreg különböző szakaszai között. A negyedik réteg azokon a területeken a legfejlettebb, ahová az afferens információ érkezik (érzékeny központok, különösen a posztcentrális gyrus). Az ötödik réteg a kéreg motoros területein fejeződik ki: precentralis gyrus, pericentralis lebeny, supramarginalis gyrus.
Minden piramis neuronnak van egy piramissejtje és sok dendritje. A piramis neuron axonja a sejt alján lévő kis dombból emelkedik ki. A kisméretű, úgynevezett interkaláris vagy intermedier piramis neuronokban a vízszintes vagy függőleges irányban elágazó axonok azonnal véget érnek, anélkül, hogy elhagynák a kéreget. A közepes és nagy piramisok axonjai sok kollaterálist adnak le a kéregben, és a fő törzsek a kéreg alatti fehérállományba kerülnek. Egy részük a kéreg alatti anyagból visszatér egy adott félteke kéregébe, vagy a corpus callosumon áthaladva a másik félteke kéregében végződik. Az agykéreg különböző részeinek egyesítésére szolgálnak. Ezért az ilyen piramis neuronokat ún asszociációs. Más axonok a kéreg alatti képződményekhez, majd az agy és a gerincvelő különböző részeihez irányulnak. Ezeket a piramisokat ún kivetítés. Az emlősök filogenetikai fejlődésével összefüggésben a piramissejtek száma nagymértékben megnő.
A teljes kéreg szélessége, a sejtek száma és az egyes sejtrétegek szélessége a frontális szakaszban, valamint a sejtösszetétel, azaz a sejtek mérete, alakja és elrendezése a kéreg egyes szakaszaiban rendkívül eltérő. . Ezek a változatok bizonyos, egyértelműen körülhatárolt területeket foglalnak el. A nagy emlősagyban 11 jól látható nagy kérgi mező található.
Szövettani és fiziológiai vizsgálatok alapján minden ilyen területet több független szakaszra osztottak. A kéreg jelentősebb differenciálódása magasabbrendű emlősöknél - majmoknál figyelhető meg. Emberben különösen nagyszerű.
Az ilyen kutatásokat Vlagyimir Alekszejevics Bets kijevi tudós indította el, aki 1874-ben publikálta a „Két központ az agykéregben” című cikket, amelyben leírta a gyrus precentrális motorzónáját és a posztcentrális gyrus érzékeny zónáját.
1909-ben Korbinian Brodmann német neurológus citoarchitektonikus mezők térképeit publikálta. agykérget. Brodmann volt az első, aki elkészítette a kéreg térképét. Ezt követően O. Vogt és C. Vogt (1919-1920) a rostszerkezetet figyelembe véve 150 mieloarchitektonikus területet írt le az agykéregben. A Szovjetunió Orvostudományi Akadémia Agyintézetében I. N. Filippov és S. A. Sarkisov az agykéreg térképét készítette, amely 47 citoarchitektonikus mezőt tartalmazott.
Adat kísérleti kutatás azt jelzik, hogy amikor az agykéreg bizonyos területeit elpusztítják vagy eltávolítják állatokban, bizonyos funkciók megzavaródnak. Ezek a tények beigazolódnak klinikai megfigyelések daganatos elváltozásokkal vagy az agykéreg bizonyos területeinek sérüléseivel rendelkező betegek számára. Mindez arra a következtetésre jutott, hogy az agykéregben vannak olyan központok, amelyek bizonyos funkciók teljesítményét szabályozzák. A fiziológiai és klinikai adatok morfológiai igazolása az agykéreg különböző részeinek szerkezetének eltérő minőségére vonatkozó doktrína volt - a kéreg cito- és mieloarchitektonikája. Megállapítást nyert, hogy a neuronok nem diffúz módon helyezkednek el a kéregben, hanem együttesekbe csoportosulnak.
A kortikális neuronok funkcióinak tanulmányozására alkalmazott modern mikroelektródos módszerek jelentősen kibővítették az érzékszervi információ feldolgozásának megértését a neokortexben és a kéreg szerkezeti szerveződését. 1957-ben V. Mountcastle amerikai kutató egy macska szomatoszenzoros (szenzomotoros) kéregében lévő sejteknek a különböző modalitású ingerekre adott válaszait elemezve a következőket fedezte fel. Érdekes tény. Amikor a mikroelektródát a szomatoszenzoros kéreg felületére merőlegesen merítették, minden sejt, amellyel találkozott, egyformán reagált egy ingerre, például a bőr enyhe érintésére vagy az ízületben történő mozgásra. Ha az elektródát a kéreg felületéhez képest szögben bemerítették, akkor az út mentén különböző szenzoros modalitással rendelkező neuronok érkeztek, bizonyos periodikussággal váltakozva.
E kísérleti tények alapján W. Mountcastle arra a következtetésre jutott, hogy a szomatoszenzoros kéreg elemi funkcionális egységek - oszlopok a felületre merőlegesen orientált. Egy ilyen oszlop átmérője körülbelül 500 μm, amelyet az afferens thalamocorticalis rost terminálisainak vízszintes eloszlása és a dendritek függőleges orientációja határoz meg. piramissejtek. Mountcastle szerint az oszlop az a szenzomotoros kéreg elemi blokkja, ahol az egyik modalitás receptoraitól származó információ helyi feldolgozása történik. A neokortex oszlopos szerveződésének ez a hipotézise széles körben elterjedt, és lendületet adott a terület további kutatásainak hazánkban és külföldön.
Alapján modern ötletek, a szenzomotoros kéreg minden funkcionális oszlopa több morfológiai mikromodulból áll, amelyek öt-hat fészek alakú neuront egyesítenek. Ez a modul több piramissejtet tartalmaz, amelyek csúcsi dendritjei a lehető legközelebb vannak, és dendrites köteget alkotnak; ezen a sugáron belül lehetséges elektrotonikus kapcsolatok, amelyek minden valószínűség szerint biztosítják a teljes egyesület szinkron működését.
A függőlegesen orientált piramissejtek csoportjával szomszédos csillagsejtek, amellyel a mikromodulhoz jutó talamokortikális rostok érintkeznek. A gátló funkciót ellátó csillagsejtek egy részének hosszú axonjai vannak, amelyek vízszintesen nyúlnak el. A piramissejtek axonjai visszatérő kollaterálisokat képeznek, amelyek egy mikromodulon belül elősegítő hatást és a mikromodulok közötti gátló interakciókat, gátló interneuronokkal érintkezve biztosíthatnak. Számos szerkezeti mikromodul, amelyet a specifikus thalamocorticalis afferensek terminálisainak vízszintes elágazása, a terminálisok axonjai és a csillagsejtek folyamatai egyesítenek, egy oszlopot (vagy makromodult) alkotnak, amelyek átmérője eléri az 500-1000 µm-t. Mindegyik oszlopra jellemző a funkcionális egység, ami abban nyilvánul meg, hogy az oszlop neuronjai egy modalitású ingerre reagálnak.
Ezt követően az oszlopos rendszerezés elvét az egyéb tanulmányozása során is megerősítették vetítési zónák ugat.
I.P. Pavlov az agykérget folyamatos érzékelési felületnek, az analizátorok kérgi végeinek gyűjteményének tekintette. Megmutatta, hogy az analizátorok kérgi vége nem valami szigorúan meghatározott zóna. Az agykéreg egy magra és szétszórt elemekre oszlik. A sejtmag a kérgi idegsejtek koncentrációjának helye, amely egy specifikus perifériás receptor összes elemének pontos vetületét alkotja, ahol a funkciók magasabb szintű elemzése, szintézise és integrációja történik. A szórt elemek mind a mag perifériáján, mind attól jelentős távolságra helyezkedhetnek el. Egyszerűbb elemzést és szintézist végeznek. A szórt elemek jelenléte a mag roncsolása során részben lehetővé teszi a károsodott működés kompenzálását. Elszórt elemek által elfoglalt területek különféle analizátorok, egymásra rétegezhetők, átfedik egymást. Így az agykéreg sematikusan elképzelhető különböző analizátorok magjainak halmazaként, amelyek között elszórtan helyezkednek el a különböző (szomszédos) analizátorokhoz tartozó elemek. A fentiek lehetővé teszik, hogy beszéljünk a funkciók dinamikus lokalizációja az agykéregben (I.P. Pavlov).
Vizsgáljuk meg a különböző analizátorok (magok) egyes kérgi végének helyzetét az ember agyféltekék gyrusaihoz és lebenyeihez viszonyítva a citoarchitektonikus térképeknek megfelelően.
1. A posztcentrális gyrus és a felső parietális lebeny kéregében idegsejtek találhatók, amelyek az általános corticalis analizátor magját alkotják. érzékenység(hőmérséklet, fájdalom, tapintás) és proprioceptív. Az agykéregbe vezető szenzoros pályák vagy a gerincvelő különböző szegmenseinek szintjén (fájdalom, hőmérséklet-érzékenység, érintés és nyomás), vagy a medulla oblongata szintjén (a kérgi irány proprioceptív érzékenységének pályái) metszik egymást. Ennek eredményeként az egyes féltekék posztcentrális gyriusai a test ellenkező feléhez kapcsolódnak. A posztcentrális gyrusban az emberi test különböző részeinek összes receptormezője kivetül oly módon, hogy az alsó testrészek érzékenységelemzőjének kérgi végei, ill. alsó végtagok, valamint a test és a fej felső részének, valamint a felső végtagok receptormezői a legmélyebben (közelebb a laterális barázdához) vetülnek.
2. Mag motor Az analizátor főként az úgynevezett motoros kéregben található, amely magában foglalja a precentrális gyrust és a paracentrális lebenyet. mediális felület féltekék. A precentralis gyrus kéregének 5. rétegében fekszik piramis neuronok (Betz-sejtek), amelyet I. P. Pavlov interkalárisnak minősített, és megjegyezte, hogy ezek a sejtek folyamataikkal kapcsolatban állnak a kéreg alatti magokkal, a koponyamagok motoros sejtjeivel és gerincvelői idegek. Ezenkívül a precentrális gyrus felső részében és a paracentrális lebenyben olyan sejtek vannak, amelyekből impulzusokat küldenek a törzs legalsó részének és az alsó végtagok izmaiba. A precentrális gyrus alsó részén olyan motoros központok találhatók, amelyek szabályozzák az arcizmok aktivitását.
Így az emberi test minden területe „fejjel lefelé” vetül a precentrális gyrusban. Tekintettel arra, hogy a gigantopiramissejtekből kiinduló piramispályák vagy az agytörzs szintjén (kortikonukleáris rostok), a gerincvelő határán, vagy a gerincvelő szakaszaiban (corticospinalis traktus) metszik egymást, a motor az egyes féltekék területei a test ellenkező oldalának vázizomzatához kapcsolódnak. Ha a végtagok izmai elszigetelten kapcsolódnak valamelyik féltekéhez, akkor a törzs, a gége és a garat izmai mindkét félteke motoros területéhez kapcsolódnak.
Motor I. P. Pavlov az agykéreg területét receptorterületnek is nevezte, mivel itt a proprioceptív (a beágyazott receptorok által érzékelt kinetikus ingerek) elemzése történik. vázizmok, inak, fascia és ízületi kapszulák).
3. A funkciót biztosító elemző mag a fej és a szemek kombinált forgatása az ellenkező oldalt , van hátsó régiók középső frontális gyrus, az úgynevezett premotoros zónában.
4. Az inferior parietalis lebeny régiójában, a gyrus supramarginalisban található a motoros analizátor magja, melynek funkcionális jelentősége a megvalósítás. szintézis mindenki céltudatos, szakszerű, munkaügyi és sportmozgalmak. Ez a mag aszimmetrikus. A jobbkezeseknél a balkezeseknél, a balkezeseknél- csak a jobb agyféltekében. Ezen összetett célirányos mozgások koordinálására való képességet az egyén élete során gyakorlati tevékenység és tapasztalatgyűjtés eredményeként sajátítja el. A célzott mozgások megvalósítása a precentrális és a szupramarginális gyrisben elhelyezkedő sejtek közötti átmeneti kapcsolatok kialakításának köszönhető. A mező károsodása nem okoz bénulást, csak az összetett, összehangolt, célirányos mozgások végzésének képességének elvesztéséhez vezet - apraxia (praxis - gyakorlat).
5. A felső parietális lebeny kéregében található az egyik sajátos érzékenységi típus bőranalizátorának magja, amelynek saját funkciója van. tárgyak felismerése érintéssel,- streognózia. Ennek az analizátornak a kérgi vége a jobb féltekében található. A kéreg felületi rétegeinek károsodása ebben a szakaszban a tárgyak érintéssel történő felismerésének funkciójának elvesztésével jár, bár az általános érzékenység más típusai megmaradnak.
6. A laterális barázda mélyén, a felső temporális gyrus insula felé eső középső részének felszínén (ahol a haránt temporális gyrus, vagy Heschl gyrus látható) egy mag található. auditív elemző. NAK NEK idegsejtek, amely a magot alkotja halláselemző a féltekék mindegyike, vezető pályák a receptoroktól mind a bal, mind a jobb oldal. Ebben a tekintetben ennek a magnak az egyoldalú károsodása nem okozza a hangok észlelésének képességének teljes elvesztését. A kétoldali károsodást kortikális süketség kíséri.
7. Mag vizuális Az analizátor található nyakszirti lebeny agyféltekék a calcarine sulcus kerületében. Mag vizuális elemző a jobb féltekét utak kötik össze a jobb szem retinájának laterális felével és a bal szem retinájának mediális felével. A bal félteke occipitalis lebenyének kéregében a bal szem retinájának laterális felének, a jobb szem retinájának mediális felének receptorai, ill. A hallóelemző maghoz hasonlóan csak a vizuális analizátor magjainak kétoldali károsodása vezet teljes kortikális vaksághoz. A vizuális analizátor magja felett kissé elhelyezkedő mező károsodása a vizuális memória elvesztésével jár, de a látás elvesztése nem figyelhető meg. Még magasabban van a kéreg egy része, amelynek veresége az ismeretlen környezetben való tájékozódási képesség elvesztésével jár.
8. Az agyfélteke halántéklebenyének alsó felületén az uncus és részben a hippocampus területén mag található szaglószervi elemző. Filogenetikai szempontból ezek a területek az agykéreg legősibb részeihez tartoznak. A szaglás és ízérzékelés szorosan összefügg egymással, ami a szagló- és ízelemzők magjainak közeli elhelyezkedésével magyarázható. Azt is megjegyezték (V.M. Bekhterev), hogy az ízérzékelés károsodik, ha a posztcentrális gyrus legalsó részének kérge károsodik. Mindkét félteke íz- és szaglóelemzőjének magjai a test bal és jobb oldalán található receptorokhoz kapcsolódnak.
Egyes analizátorok leírt kérgi végei nincsenek jelen az agykéregben csak az emberek, hanem az állatok is. A külső és belső környezetből érkező jelek észlelésére, elemzésére és szintézisére specializálódtak, amelyek I. P. Pavlov meghatározása szerint a valóság első jelrendszerét alkotják.
Második jelzőrendszer csak az emberekben létezik, és a beszéd fejlődése határozza meg. A beszéd és vele együtt a tudat filogenetikailag az agy legfiatalabb funkciója. Ebben a tekintetben az analizátorok kortikális végei a legkevésbé lokalizáltak. A beszéd- és gondolkodási funkciókat a teljes kéreg részvételével hajtják végre. Az agykéregben azonban meg lehet különböztetni bizonyos területeket, amelyeket szigorúan meghatározott beszédfunkciók jellemeznek. Igen, elemzők motoros beszéd(szóbeli és írásbeli) a kéreg motoros területe mellett találhatók, pontosabban a frontális lebeny kéregének azon részein, amelyek szomszédosak a precentrális gyrusszal.
Elemzők vizuális és auditív észlelés beszédjelek a látás- és halláselemzők mellett helyezkednek el. Ki kell emelni, hogy a jobbkezesek beszédelemzői a bal féltekében, a balkezeseknél pedig főleg a jobb féltekében találhatók.
Nézzük meg néhány beszédelemző helyzetét az agykéregben.
1. Az írott beszéd motoros elemzője a középső frontális gyrus hátsó részén található. Ennek az elemzőnek a tevékenysége szorosan összefügg a céltudatos mozgások elemzőjével, amely biztosítja az íráshoz szükséges tanult kéz- és szemmozgások kialakítását. Sérülés esetén a finom mozgások végzésének képessége, amelyet általában a látás ellenőrzése alatt hajtanak végre, és a betűk, szavak és egyéb jelek rajzolásához szükséges, elveszik (agraphia).
2. Motorelemző mag beszéd artikuláció(beszédmotoros analizátor) a gyrus inferior frontális részében (Broca központja) található. A kéreg károsodása ezen a területen motoros afáziához, azaz a szavak kiejtésének képességének elvesztéséhez vezet (afázia). Ez az afázia nem jár együtt a beszédtermelésben részt vevő izmok összehúzódási képességének elvesztésével. Sőt, ha ez a mező megsérül, a hangok kiejtésének vagy az éneklés képessége nem vész el.
3. B központi osztályok inferior frontális gyrus található az énekléshez kapcsolódó beszédelemző magja. Vereségét vokális amuzia kíséri – képtelenség zenei frázisok komponálására és reprodukálására, valamint agrammatizmus, amikor az egyes szavakból értelmes mondatok összeállításának képessége elvész. Az ilyen emberek beszéde olyan szavak halmazából áll, amelyek szemantikai értelemben nem kapcsolódnak egymáshoz.
4. Mag auditív beszédelemző szorosan összefügg
a hallóanalizátor kérgi központja, és a hátsó szakaszokban található
felső temporális gyrus, a felületén oldalra néz
az agyfélteke laterális barázdája. A mag károsodása általában nem rontja a hangok hallási észlelését, de a szavak és a beszéd megértésének képessége elvész (verbális süketség, vagy szenzoros afázia). Ennek a magnak az a funkciója, hogy az ember nemcsak hallja és megérti egy másik személy beszédét, hanem irányítja a sajátját is.
5. A felső temporális gyrus középső harmadában található a corticalis analizátor magja, melynek veresége a zenei süketség kialakulásával jár együtt, amikor a zenei frázisokat különféle zajok értelmetlen halmazaként érzékeljük. A halláselemzőnek ez a kérgi vége a második jelrendszer központjaihoz tartozik, amelyek érzékelik a tárgyak, cselekvések, jelenségek verbális kijelölését, pl. jelérzékelő jelek.
6. A vizuális analizátor magjával közvetlen kapcsolatban van egy mag az írott beszéd vizuális elemzője(39. mező), az alsó parietális lebeny szögletes gyrusában található. Ennek a magnak a károsodása az írott szöveg észlelésének és olvasási képességének elvesztéséhez vezet (diszlexia).
Az emberi frontális kéreg a társadalmi elvhez kapcsolódik. Neuronjai kétirányú kapcsolatot alakítanak ki a kéreg összes többi területével. Károsodás esetén rendellenességek alakulnak ki társadalmi viselkedés, durvaság, hűtlenség, csökkent céltudatosság, apátia. A lobotómia után teljesen hiányzik a kezdeményezőkészség és a társadalmi aktivitás.
A jobb és bal agyfélteke funkcióinak aszimmetriája nagyon hangsúlyos. A bal agyfélteke biztosítja a logikát, az elvont gondolkodást, dominál a beszédfunkciók (olvasás, írás, számolás), valamint a bonyolult akaratlagos mozgások vonatkozásában. Jobb agyfélteke– emocionalitás, imaginatív gondolkodás, összetett vizuális és auditív képek felismerésének beszéd nélküli funkciói, tapintható érzékelés, tér-, forma-, irányérzékelés, intuitív gondolkodás.
Hasonló cikkek
-
Csernisov: Nem érdekelnek azok a képviselők, akik nevettek rajtam!
Borisz Csernisov, az LDPR Állami Duma képviselője az egyik legfiatalabb parlamenti képviselő. 25 éves. Valamivel több mint három hónapig dolgozott az alsóházban – és már két elpárologtatás elleni törvényjavaslatot is benyújtott. ViVA la Cloud nyílt forráskódon...
-
Az LDPR-helyettes megcsalta a feleségét, megfenyegette a szeretőjét, megverték és önmagára támadt
Pályafutását 1986-ban kezdte a Trud újság kiadójánál, 18 évesen behívták a hadseregbe katonai szolgálatra, amelyet 1987-től 1989-ig teljesített. Krasznojarszk Az 1990-es évek végén a krasznojarszki kormányzói hivatalban kapott állást...
-
Ha belépsz a Liberális Pártba, az mit ad neked?
Bevezetés………………….………………...………….……. 3 1. fejezet. Az LDPR jogalkotási tevékenysége a Dumában ..... 8 2. fejezet A frakciótagok munkája az Állami Duma bizottságaiban ......... 10 Következtetés .......... .................................................. .......... ... 13 Források és irodalom jegyzéke ………………………….. 14...
-
A kalapok visszaválthatók?
Ha 2019-ben azon töprengett, hogy a vásárlás után visszaküldhető-e egy kalap az eladónak (üzletnek vagy magánszemélynek), és pénzt kaphat - olvassa el a cikket, és megtudja, milyen esetekben és hogyan lehet kalapot visszaküldeni. ..
-
"Atommag fizika és technológia" szak (alapképzés)
Korábban ennek az állami szabványnak a száma 010400 (a felsőoktatási szakoktatási irányok és szakok osztályozója szerint) 4. A főképzési program tartalmi követelményei OKTATÁSI MINISZTÉRIUM...
-
Az oroszországi rendkívüli helyzetek minisztériumának felsőoktatási intézményei
A Polgári Védelmi Akadémia fogadja azokat az állampolgárokat, akik államilag kibocsátott középfokú (teljes) általános vagy középfokú szakképzésről szóló okirattal, valamint alapfokú szakképzési oklevéllel rendelkeznek, ha az rekordot tartalmaz...