Od čega zavisi skelet parnih udova sisara? Vanjska struktura sisara, skeleta i mišića na primjeru domaćeg psa
Jedna od karakteristika skeleta sisara je veliki razvoj hrskavice kod mladih životinja i njeno dugoročno očuvanje u postembrionalnom periodu razvoja. To se objašnjava karakteristikama rasta hrskavice i kostiju u vezi s rastom same životinje. To je hrskavica koja raste cijelom svojom masom (intersticijsko), rast se odvija brzo i proporcije dijelova se mogu održati, dok kost raste sporije i samo s površine, tako da dijelovi skeleta koji se pojavljuju uvijek ostaju na istom rastojanju i proporcije delova su očuvane tokom rasta kosti nemoguće. Zbog toga mlade životinje uvijek sadrže mnogo hrskavice, a posebno na mjestima gdje je rast intenzivan. Dakle, duge kosti udova uvijek se sastoje od srednjeg dijela - dijafize, koja prvo okoštava, i krajeva koji dugo ostaju hrskavičasti - epifize. Osifikacija dijafize počinje prvenstveno od periferije - perihondralne, tj. zbog ćelija koje potiču iz vezivno tkivo, koji pokriva hrskavicu, - od perihondrija; zatim mu se pridružuje unutrašnje okoštavanje hrskavice - endohondralno okoštavanje; tek kasnije, kada se u dijafizi pojave šupljine zbog sloma kostiju koštana srž, epifize počinju endohondralno okoštavati; Između dijafize i epifize, međutim, dugo ostaje hrskavica, uz pomoć koje kost raste u dužinu uz zadržavanje proporcija (Sl. 530).

Kičma sisare, za razliku od Sauropside, karakterizira odsustvo slobodnih artikulacija između pršljenova, s izuzetkom prvog i drugog pršljena. Intervertebralni elastični diskovi vezivnog tkiva nalaze se između tijela pršljenova. Općenito, to rezultira manjom fleksibilnošću kralježnice, što je posljedica specijalizovanijeg kretanja sisara uz pomoć udova.
Okoštavanje tijela pršljenova događa se kroz hrskavične epifize. Tijela imaju ili ravne površine ili blago konveksne, a kod većine modernih kopitara blago su opistocela. Dobro razvijen na pršljenima, posebno u područje grudi, spinozni nastavci, kao i poprečni nastavci koji sjede na lukovima; u osnovi potonjeg nalaze se zglobni procesi.

Broj vratnih pršljenova je obično 7 (sl. 531), bez obzira na dužinu vrata, tako da dužina vrata, za razliku od ptica, zavisi isključivo od dužine pršljenova. Samo lamantin (Manatus) i jedna vrsta lenjivca (Choloepus hoffmani) imaju 6 vratnih pršljenova, dok troprsti lenjivci (Bradypus) imaju 9. Skoro izumrli zbog vodenog načina života, vrat kitova još uvek sadrži 7 ravnih, delimično sraslih pršljenova, dok je u vratu žirafe ovih 7 pršljenova veoma dugačko. Prva 2 vratna pršljena su modificirana u atlas i epistrofiju, povezani pravim zglobovima, iu monotreme odontoidni proces još uvijek funkcionira u obliku posebne kosti i nije srasla, kao kod drugih sisara, s tijelom epistrofeja. Rudimentarna rebra cervikalne regije stapaju se sa pršljenom, što rezultira stvaranjem perforiranih poprečnih procesa. Djelomično ili potpuno spajanje vratnih pršljenova javlja se kod kitova; kod jerboa se spajaju svi vratni pršljenovi osim 7., u tobolastom madežu (Notoryctes) spajaju se 2. do 6. pršljen, u mladežu - od 3. do 4.
Broj torakalnih pršljenova, za koje je karakteristično vezivanje rebara za njih, varira od 9 [kod jednog kitova (Hyperoodon) i armadila (Tatusia)] do 25 [kod ljenjivca], ali obično ih ima 12-13 . Torakalni pršljenovi imaju dobro razvijene procese: spinozne (proc. spinosus), poprečne ili dijapofize (proc. transversus), zglobne ili zigapofize (proc. articularis); Rebra su svojim glavama spojena sa tijelom jednog ili dva susjedna pršljena, a svojim tuberkulima sa poprečnim nastavcima. Sastoje se od dva dijela: koštanog rebra - proksimalnog i hrskavičnog rebra - distalnog. Samo rebra prednjih torakalnih pršljenova dopiru do prsne kosti, stražnja rebra završavaju slobodno ( lažna rebra). Lumbalni pršljenovi, koji se odlikuju odsustvom rebara koja su ovdje uključena u poprečne procese, kod sisara se kreću od 2 do 9, obično ih ima 6-7.
Sakralni pršljenovi sisara su oni pršljenovi koji, spojeni zajedno, artikuliraju sa ilium. Takvih pršljenova ima 1-10, ali samo 1-2 od njih su pravi sakralni pršljenovi, odnosno oni kod kojih se artikulacija sa karlicom odvija kroz nezavisne osifikacije koje odgovaraju sakralnim rebrima; preostali sakralni pršljenovi su lažni, tj. kaudalni pršljenovi spojeni sa sakrumom.
Broj pršljenova uključenih u kaudalnu regiju veoma varira: od 3 (šišmiši, giboni) do 49 [gušter (Manis macrura)].
Grudna kost(sternum) sisara nastaje spajanjem trbušnih krajeva rebara, koji zatim formiraju jednu nesparenu ploču, koja se sastoji od više dijelova (Sl. 532).

Spojen na ključne kosti (ako postoje) i na prvi par rebara, proširen prednji dio grudna kost se naziva manubrijum (praesternum, manubrium); Slijedi dio koji se naziva tijelo (mesosternum, corpus), koji se često razbija u niz kostiju, sukcesivno smještenih između uzastopnih parova rebara; još pozadi leži tzv xiphoid process(xiphisternum). Nisu sva rebra povezana sa grudne kosti, manje ili više značajan broj lažnih rebara završava slobodno (Sl. 533).

Kod kloakalnih sisara (Monotremata), zbog prisustva korakoida, kojih nema kod svih ostalih sisara, predsternalna hrskavica (prosternum) je položena ispred manubrijuma grudne kosti, koji je u vezi sa korakoidima, koji je tada zamijenjena kostom u razvoju - episternumom, koja kloakalne sisare približava gmizavcima.
Scull Sisavci imaju niz osobina koje se razlikuju od lubanja reptila. Međutim, ove osobine nisu se pojavile odjednom, već kroz postepene promjene, što se može vidjeti proučavanjem lubanja fosilnih guštera (Theromorpha), posebno iz reda Theriodontia. Lobanja sisara je tropibazalna lobanja, ali sa skraćenim interorbitalnim dijelom, zbog čega izgleda kao platibazalna, sa velikom lobanjom, sa jasnom tendencijom spajanja pojedinačnih kostiju,
Raspored kostiju u lobanji sisara može se suditi iz priloženog dijagrama (sl. 534).
Na bočnim okcipitalnim kostima, prema van od zglobnih tuberkula, obično se nalazi okcipitalni nastavak (proc. paroccipitalis), posebno razvijen kod glodara i mnogih kopitara. Ispred zglobnih tuberkula obično postoji otvor za izlaz hipoglosalnog (XII) živca (for. condyloideum anterius).
U hrskavici slušne čahure položeno je nekoliko okoštavanja, koja se, međutim, vrlo rano spajaju u dvije kosti: onu koja leži ispred - petrosum - i onu koja leži spolja i iza - mastoid (mastoideum); obje ove kosti, spojene zajedno, stvaraju petromastoidnu, ili jednostavno petroznu kost (petromastoideum, petrosum ili perioticum), koja zatvara unutrašnji slušni labirint. Periotikum se graniči pozadi sa glavnim i bočnim okcipitalne kosti, a u prednjem dijelu sa sfenoidnom i pterigosfenoidnom kosti; budući da sudovi i živci prolaze između ovih kostiju i petrozne kosti, ovdje ostaju značajne praznine: pozadi - stražnji raščupani foramen (for. lacerum posterius), ili jugularni foramen (for. jugulare) za prolaz glosofaringealnog (IX ) nerv, grane vagusnog (X) nerva i za pomoćni (XI) nerv, kao i za unutrašnji jugularne vene, a ispred - prednja neravnina (for. lacerum anterius), kroz koju prolazi unutrašnja karotidna arterija. Ponekad je veza između petrozne kosti i lubanje toliko slaba da se lako može odvojiti zajedno sa kosti koja je za nju pričvršćena. bubna kost ili čak tokom maceracije lobanje potpuno ispadne, kao što je slučaj kod kitova.
Izvan ušne regije formiraju se dvije integumentarne kosti. Jedan od njih, čije porijeklo još uvijek nije dovoljno razjašnjeno, razvija se u oblasti eksternog ušni kanal u obliku polukružne male kosti. Ovo je bubna kost (tympanicum), koja je vjerovatno homolog ugaone kosti (angulare) donje čeljusti gmizavaca, koja se preselila u parotidnu regiju zajedno sa ostalim kostima donje čeljusti zbog promjena koje su se dogodile na čeljusti artikulacija. U početku se nalazi u obliku polukružne ili prstenaste kosti u predjelu slušnog kanala, često raste i formira, najčešće zajedno s petroznom kostom, a ponekad i s ljuskavom kosti, vanjski slušni kanal (meatus auditorius externus). Postoji napetost na bubnoj kosti bubna opna. Dublje unutra, bubna kost takođe može učestvovati u formiranju zidova srednjeg uha. Često bubna kost na donjoj strani slušne regije nabubri u mehur tankih zidova (bulla ossea).
Druga kost dermalnog porijekla koja učestvuje u formiranju skeleta ušne regije je ljuskava kost (squamosum); dobila je izuzetan razvoj i značaj kod sisara zbog pričvršćenja donje vilice za nju. Kod majmuna i ljudi, sve ove kosti regije uha su spojene zajedno da formiraju složenu temporalnu kost (temporale); Ljuskava kost ovdje čini samo dio, odnosno ljuske (squama temporalis) temporalna kost, dok je bubna kost bubanj dio (pars tympanicum), a pravi parotidni hrskavične kosti formiraju kameni (pars petrosa) i mastoidni (pars mastoidea) dio. Stepen razvijenosti skvamozne kosti i njeno učešće u formiranju zidova moždanog omotača i slušnog kanala su veoma različiti, ali pločasta kost uvek ima zglobnu površinu (cavitas glenoidea) za artikulaciju donje čeljusti. Dakle, kod sisara, za razliku od svih drugih kralježnjaka, postoji vrlo posebna originalna artikulacija donje čeljusti: ne na četvrtastu kost, već na skvamoznu kost lubanje. Zglobna površina donje vilice leži na bazi zigomatskog nastavka (proc. zygomaticus) koji se proteže prema van i naprijed od ljuski. Kod monotremesa, iznad zglobne površine, probijen je temporalnim kanalom (can. temporalis), posebno dobro izražen u platipusu. Iza zglobne jame nalazi se proces (proc. postglenoideus).
Ispred slušne regije, baza lubanje je formirana od glavnog sfenoidna kost(basisphenoidale), na čijoj se unutrašnjoj površini nalazi manje ili više značajno udubljenje - tzv. turcica sela (sella turtica), u kojoj se nalazi hipofiza. Sa strane glavne sfenoidne kosti leže, tvoreći donje dijelove prednjeg zida lubanje, pterigosfenoidne kosti (alisphenoidea), obično srasle s njom u jednu kost, tvoreći velika krila (alae magnae) ove kosti. Pterigosfenoidne kosti opremljene su sa dva otvora: ovalnim (for. ovale) za prolaz treće grane trigeminalni nerv(V3) i okrugli (for. rotundum) za prolaz druge grane istog živca (V2). Međutim, prvi se često spaja sa prednjim napuknutim foramenom (for. lacerum anteris), a drugi se često spaja sa sfenorbitalnim foramenom (for. sphenorbitale), koji je istovremeno omeđen i pterygosfenoidnom kosti i orbitalno-sfenoidnom kosti koja leži ispred to. Kroz ovaj otvor prolaze okulomotori (oculomotorius III), trohlearni (trochlearis IV), abducens (VI) i prva grana trigeminalnog živca.
Lobanja je ovdje prekrivena tjemenim kostima (parietale), koje često srastaju (kod monotremesa, nekih torbara i kopitara); između njih i gornje okcipitalne kosti položene su još dvije kosti koje se spajaju u jednu međuparijetalnu kost (interparietale), koja je u nekim redovima očuvana kao samostalna kost (kitovi, tobolčari, hiraksi, aardvarci, vunasta krila i neki glodari), dok u drugima se spaja ili s parijetalnom (kod nekih glodara, sirena i preživara), ili s gornjom potiljnom kosti (kod mesojeda, jednopapka, primata).
Ispred glavne sfenoidne kosti leži prednja sfenoidna kost (praesphenoideum) sa udubljenjem za hijazmu oftalmoloških nerava; na njegovim stranama, obično srasle s njim, leže orbitalno-sfenoidne kosti (orbitosphenoidea) s otvorom za prolaz orbitalnih živaca (for. opticum), koje se nazivaju mala krila (alae parvae) u slučaju spajanja sa prednje i glavne sfenoidne kosti. Kao što je već spomenuto, između obje krilaste kosti sa svake strane postoje otvori u obliku proreza kroz koje prolaze živci očne mišiće i prva grana trigeminalnog živca (for. sphenorbitale).
Na vrhu ovog reda kostiju odgovaraju integumentarne prednje kosti (frontale) - obično dobro razvijene parne kosti, ponekad srasle i nose šipke kod goveda, i panjeve za rogove kod jelena. Prekrivajući lobanju, prednje kosti su ograničene iznad i sa unutra orbite, često formiraju procese: preorbitalni (proc. praeorbitalis i postorbitalis (proc. postorbitalis). Ponekad oko očiju nastaje periorbitalni prsten zbog spajanja zigomatične kosti (jugale) sa frontalnom kosti (neki insekti i mesožderi, preživari). Kod majmuna proces odvajanja orbitalne utičnice od temporalne šupljine ide toliko daleko da se između orbite i temporalne šupljine formira koštani zid.
U prednjem zidu orbite leži mala integumentarna suzna koščica (lacrymale), perforirana otvorom suznog kanala (for. lacrymale).
U nosnoj regiji formira se nekoliko okoštavanja u hrskavičnoj nosnoj kapsuli. Prvo, duž hrskavičnog nosnog septuma, u sredini se razvija koštani septum (lamina perpendicularis), koji odgovara srednjoj olfaktornoj kosti (mesethmoideum); drugo, u zadnji zid nosne kapsule iu prednjem zidu moždane šupljine razvijaju se dvije lateralne ploče, perforirane brojnim [samo kod platipusa ove rupe su zamijenjene jednim (za. olfactorium)] otvorima za prolaz njušnog živca - kribriformnim pločama (lamina cribrosa), što odgovara bočnim etmoidalnim kostima (ethmoidalia lateralia). Od nosnog septuma, okoštavanje se širi s dorzalne strane duž bočnih strana, prolazi duž bočnih stijenki nosnih kapsula i završava na dnu, tvoreći brojne izbočine sa strana u nosnu šupljinu. To su olfaktorne školjke (ethmoturbinalia), koje zajedno sa bočnim pločama i šupljinama ćelijske strukture ponekad čine vrlo složen nosni labirint. Položaj školjki može se vidjeti iz priloženog šematskog crteža (sl. 535). Sa integumentarne nosne kosti (nasale), koja prekriva ovo područje na vrhu, školjke (nasoturbmalia) vise u nosnu šupljinu, takođe po jedna sa svake strane. Odozdo nosni septum prekriva nesparenu kost, za koju se ranije smatralo da je vomer. Međutim, nedavne studije su pokazale da se, pored ove nesparene integumentarne kosti, u bazi lubanje razvijaju dvije uparene kosti, nazvane prevomer; a budući da je kod gmizavaca vomer prisutan u obliku parne kosti, a nesparena kost na ovom području je bila parasfenoid, onda se vomer sisara može smatrati homologom ove posljednje kosti kod gmizavaca, a prevomers se može smatrati vomers. Ispred zidova nosne šupljine nalaze se kosti vilice (maxillare) na kojima se formiraju školjke koje prerastaju u nosna šupljina; ovo su ljuske vilice (maxilloturbinale).

Prednja strana lubanje se sprijeda završava premaksilarnim kostima (praemaxillare). Kod antropomorfnih majmuna i ljudi, ova kost se vrlo rano spaja s kosti vilice, a kod kiroptera je reducirana na malu kost ili čak i potpuno odsutna. Svojim okomitim procesom, premaksilarna kost ograničava nazalni otvor sprijeda i proteže se između nosne kosti i kosti čeljusti daleko nazad, ponekad do frontalne kosti. Ispod, u formiranju učestvuju premaksilarne kosti tvrdo nepce. Oni ograničavaju prednje i bočne otvore palatinskog (Stensonovog) kanala (for. incisiva) na tvrdom nepcu, povezujući usnoj šupljini sa Jacobsonovim organom. Procesi premaksilarnih kostiju koji leže izvan ovih palatonazalnih otvora su superponirani za svaku kost nezavisno, vjerovatno su homolozi uparenog vomera gmizavaca (praevomer) iu platipusu su sačuvani kao samostalne kosti. Sjekutići se nalaze na premaksilarnoj kosti.
Kosti vilice (maxillare), kao što je već spomenuto, ograničavaju dno nosne šupljine, formirajući tvrdo nepce horizontalnim procesima. Na unutrašnjoj strani nosne šupljine, donja nosiva školjka (maxilloturbinae) raste do njegovih bočnih zidova, formirana od kostiju vilice, i formirana je kao samostalna kost.
Nastavak leđa tvrdog nepca su horizontalni nastavci palatinskih kostiju (palatinum), koji ograničavaju unutrašnji nosni prolaz kao bočne stijenke. Stoje okomito, a pozadi su im spojene i okomito stojeće pterygoideum kosti, koje se kod ljudi spajaju sa sfenoidnom kosti, formirajući njene pterygoideum nastavke (proc. pterygoidei).
Prednji dio lubanje podvrgava se značajnim modifikacijama u vezi s prelaskom na vodeni način života kod kitova, sirena i u vezi s razvojem trupa kod slonova.

Kod kitova (Sl. 536), nozdrve se pomeraju daleko nazad u interorbitalni region i vode u vertikalni kanal, koji se sa hoanama otvara u usnu duplju; nosne kosti leže iza nozdrva, preklapajući čeone kosti i gotovo uz međuparijetalnu kost; Također, kosti vilice preklapaju čeone kosti, protežući se gotovo do parijetalnih kostiju, snažno pomaknute u stranu od strane interparijetalne kosti.
Kod sirena se dešava isto kretanje unazad nozdrva, ali ne tako daleko; nosne kosti su ovde još više smanjene i mogu čak i da izostanu (Sl. 537).

Kod slonova, s pretjeranim razvojem kljova i snažnih kutnjaka i zbog prisustva trupa, normalan raspored kostiju je uvelike izmijenjen: nozdrve su snažno potisnute unazad, te stoga nosni kanal zauzima gotovo okomit položaj; prednje kosti su vrlo kratke, premaksilarne kosti su vrlo velike i šire se daleko unazad; kosti vilice su pomaknute ispod baze lubanje; cijela lobanja postaje vrlo masivna sa izrazito razvijenom pneumaticnošću kostiju (sl. 538).
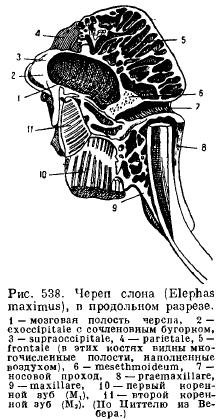
Donja čeljust sisara, za razliku od svih ostalih kralježnjaka, razvija se sa svake strane samo od jedne zubne kosti (dentale); obje polovice ili ostaju povezane jedna s drugom pomoću hrskavice, šava ili rastu zajedno (jednopapke, slonovi, slepi miševi, majmuni). Svaka polovica se sastoji od horizontalnog tijela (korpusa), na kojem se nalaze zubi, i uzlaznog dijela s nastavcima: prednjim koronoidni proces(proc. coronoideus), zglobni nastavak koji leži iza njega (proc. articularis) i kod nižih sisara nazadni ugaoni nastavak (proc. angularis); kod većine torbara i nekih glodara ovaj proces se pretvara u ploču koja se proteže prema unutra (Sl. 539).
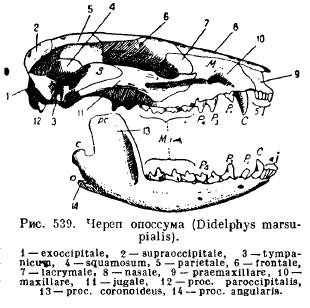
Ovakva struktura donje čeljusti i njeno osebujno vezivanje za lubanju dovode nas do pitanja porijekla ovih osobina kod sisara. Priča individualni razvoj a paleontološki podaci daju odgovor na to pitanje. U embrionima sisara formiraju se palatokvadrat i Mekelove hrskavice. Međutim, okoštavanje palatokvadratne hrskavice nastaje nezavisno od artikulacije donje čeljusti i ne dovodi do formiranja kvadratne kosti, kao kod gmizavaca, već će dovesti do nove slušne kosti - inkusa (inkusa), dok Mekelova hrskavica, okoštavajući u svom stražnjem dijelu, stvara nezglobnu kost.kost (articulare), kao i drugi gnatostomi, a treća slušna kost je malleus (malleus). Prva slušna koščica, stapes (stapes), odgovara stubu vodozemaca, gmizavaca i ptica i, kao što već znamo, predstavlja modifikovani ovjes (hyomandibulare), odnosno proksimalni dio hioidnog luka. Kao i stub (columella auris), uzengija pokriva ovalni prozor slušni labirint, a perforiran je, kao kod nekih gmizavaca, otvorom kroz koji prolazi facijalna arterija (a. facialis) kod nekih nižih sisara (sl. 540 i 541).

Tako se kod sisara kvadratna kost pretvorila u inkus, zglobna kost u malleus; a budući da mala integumentarna kost raste do malleusa, formirajući njegov srednji nastavak, vjeruje se da ovaj posljednji odgovara integumentarnoj zglobnoj kosti (goniale) reptila. Integumentarna ugaona kost gmizavaca, koja leži pored ove poslednje kosti, takođe prelazi u region uha, postajući bubna kost (tympanicum). Ove promjene vidljivo na priloženom dijagramu (sl. 541).
Paleontološki podaci potvrđuju mogućnost navedenih transformacija. Među zvjerozubim gušterima (Theriodontia) postoje oni kod kojih zubna kost dobiva posebno veliki razvoj, dopirući iza mjesta zgloba donje čeljusti sa kvadratnim manusom. U isto vrijeme, skvamoza se spušta duž kvadrata do iste artikulacije. Zubne i skvamozne kosti se tako sve više zbližavaju.
Konačno, kod nekih sisara u artikulaciji sudjeluju zubne i skvamozne kosti zajedno sa zglobnim i kvadratnim kostima.
Što se tiče hioidnog aparata sisara, on se sastoji od tijela, modificiranog bazalnog dijela (copula) hioidnog luka, prednjih rogova (cornua anteriora), formiranih od donjeg dijela hioidnog luka, i stražnjih rogova (cornua posteriora) , transformisan iz prvog grančičnog luka. Prednji rogovi se sastoje od nekoliko okoštavanja (do četiri); segment okrenut ka lubanji često raste na kamenu kost, formirajući stiloidni nastavak (proc. styloideus).
Drugi i treći škržni luk doveli su do formacije tiroidna hrskavica(cartilago thyreoidea.)
Pojasevi za ekstremitete. Rameni pojas sisara karakteriše prvenstveno redukcija kostiju gavrana (procoracoideum, epicoracoideum, coracoideum), težnja ka nestanku klavikule (clavicula) i dobro razvijenim grebenom lopatice (spina scapulae), nastavljajući se u akromion proces (akromion).
Međutim, kod monotreme rameni pojas je vrlo sličan reptilima. Lopatica je opremljena akromijalnom izbočinom na koju se sprijeda prislanja klavikula, a pozadi su za lopaticu pričvršćene široke i kratke gavranove kosti (coracoideum) koje idu do prsne kosti. Ispred njih se nalaze široki prokorakoidi (procoracoideum) u obliku nezavisnih kostiju. Ključne kosti (klavikula) se spajaju medijalno sa episternumom (slika 542). Kod drugih sisara, krakoidi se formiraju u obliku dva okoštavanja, ali gube vezu sa sternumom i rastu do lopatice, formirajući korakoidni nastavak (proc. coracoideus); prokorakoidi potpuno nestaju (Sl. 543), klavikule nestaju kod nekih mesoždera i kopitara zbog prilagođavanja na brzo trčanje, a kod kitova i sirena zbog transformacije prednjih udova u peraje.


Zdjelični pojas sisara ima tipičan raspored elemenata kada se formira; njegova posebnost je, prvo, prisustvo dodatne kosti u acetabulumu (acetabulare), drugo, spajanje svih ovih kostiju sa svake strane u jednu bezimenu kost (innominatum) i fuzija pubične i ishijalne kosti, tako da zatvorena struktura formirana sa svih strana postoji rupa između ovih kostiju (for. obturatum) (sl. 544). Kod monotremesa i tobolčara, posebna kost je pričvršćena za stidnu kost odozdo kako bi poduprla burzu (marsupiale).
Skelet ekstremiteta kod izvornih oblika sisara je zbog njihove adaptacije na brze pokrete – trčanje uz pomoć četiri uda, koji se nalaze ispod tijela, tako da potonji ne visi između udova, kao kod većine gmizavaca, već se oslanja direktno na njima. Sposobnost trčanja povezana je s različitim položajem udova kod sisara. Njihovi su udovi orijentirani tako da su laktovi prednjih udova usmjereni prema nazad, a zglobovi koljena stražnjih udova usmjereni prema naprijed. Prilikom kretanja, prednji udovi se zabacuju naprijed i povlače tijelo savijanjem zglobova ručnih i laktova; Zadnji udovi guraju tijelo naprijed ispravljajući nogu u zglobu koljena.

U pogledu broja kostiju uključenih u udove i njihove lokacije, skelet udova predačkih oblika sisara ima vrlo tipičnu strukturu primitivnog petoprstog uda i doživio je velike promjene unutar klase tek u procesu. prilagođavanja različitim vrstama pokreta u različitim uslovima okruženje.
Prednji ud karakterizira prisustvo u distalnom dijelu humerus otvori (for. entepicondylare), prisustvo proksimalne izbočine ulna(olecranon), prisustvo pisiforme kosti (pisiforme) i fuzija četvrte i pete distalne karpalne kosti (carpalia dis talia 4+6 = hamatum, ili uncinatum) i centralne proksimalne s intermedijalnom (centrale dist. + intermedium = lunatum, ili semilunare). Zbog činjenice da se nomenklatura kostiju ručnog zgloba sisara, usvojena u ljudskoj anatomiji, često koristi, prikazujemo tablicu ovih naziva u dijagramu koji označava lokaciju kostiju.
Stražnji ud sisara karakteriše prvenstveno činjenica da je zglob koljena rotiran naprijed, bedro ima dobro razvijene procese na proksimalnom dijelu - trohanteri, uključujući 2-3 (trohanter veliki, manji i tertius); u potkoljenicu tibija razvijenije, tu je kost kapa za koljeno(patela), zaštita kolenski zglob. Tarsus ima samo dvije proksimalne kosti, dobro razvijene: unutrašnju talus (talus = astragalus), formiranu fuzijom srednjeg i centralnog proksimalnog (intermedium + centrale proximale), i vanjski kalkaneus (calcaneus), koji odgovara fibularnom tarzalu kost (fibulara). Od koštica srednjeg reda centralna distalna (centrale distale) prelazi u navikularnu (naviculare = scaphoideum). Tibijalna kost je ili smanjena ili spojena sa ovom potonjom.
Raspored tarzalnih i metatarzalnih kostiju i sinonimija imena je sledeća:
U vezi s prilagodbom na kretanje u različitim sredinama, ekstremiteti sisara su pretrpjeli vrlo značajne promjene, a te promjene se posebno jasno mogu pratiti na skeletu. Nažalost, nemamo paleontoloških podataka o građi udova predaka sisara. Nema sumnje da su prvi sisari imali slabo specijalizirane udove gore opisanog tipa. Među našim modernim sisavcima, najprimitivniji tobolčari, na primjer, oposumi - tobolčarski štakori (Didelphys), i insektojedi koji vode arborealni način života, imaju upravo ove udove. Međutim, iz ovoga bi bilo pogrešno zaključiti da su preci sisara zasigurno bili stanovnici šuma i, osim toga, forme koje se penju na drveće. Osim toga, oposumov ud, iako se koristi za kretanje po granama drveća, nikako nije tipičan specijalizirani arborealni ud, već je zadržao svoj univerzalni petoprsti tip. Takvi udovi mogli bi se najbolje sačuvati kod životinja koje ne samo da su trčale po zemlji, već su se pele i na drveće, kao i kod životinja koje su se penjale na planine. Specijalizacija udova sisara dogodila se istovremeno sa diferencijacijom redova i drugih manjih sistematskih jedinica, a došlo je i do konvergencije.
Udovi prolaze kroz posebno duboke modifikacije kada se prilagođavaju letu, plivanju u vodi i kretanju ispod zemlje; sporo kretanje na tlu i arborealni način života nisu posljedica duboke specijalizacije.
Kod slepih miševa (Chiroptera) prednji ud je transformisan u krilo, a letna membrana je razvučena između četiri (2-b) visoko izdužena prsta bez kandži; prvi prst je kratak i na kraju ima jako razvijenu kandžu; ovaj prst ne igra nikakvu ulogu u letenju, koristi se za penjanje; u zglobu se spajaju skafoidna, lunasta i centralna kost; podlaktica je izdužena i sastoji se od dobro razvijenog radijusa, dok je ulna smanjena i prianja uz radijus. Leteća membrana pokriva i zadnje noge. Između ovih, različito je razvijen kod slepih miševa (Sl. 545).
Kod sisara koji plivaju u vodi, udovi se pretvaraju u peraje na sličan način, kao što smo već vidjeli kod gmizavaca. Udovi kitova i sirena su se posebno dramatično promijenili. Za razliku od krila, prednji ud je značajno skraćen u svom proksimalnom dijelu. Kod kitova su kosti ramena i podlaktice postale kratke i široke ploče, pružajući prostor za pričvršćivanje snažnih mišića. Svi dijelovi udova su nepomično povezani jedni s drugima, formirajući snažno veslo. U ovom slučaju, broj falangi se može znatno povećati. Videli smo iste promene kod gmizavaca. Povećanje broja falangi ovisi o činjenici da se dijafize falanga ne spajaju s epifizama i okoštavaju iz odvojenih centara. Ovo zasebno okoštavanje nastaje zbog sporijeg procesa okoštavanja kod ovih životinja. Sporost procesa okoštavanja dovodi do činjenice da većina kostiju na njihovim spojevima ostaje hrskavična.
Peraje sirena (Sirenia) su po izgledu slične perajima kitova: rame i podlaktica su zglobljeni tako da se mogu savijati; ima skoro sve kosti ručnog zgloba i svih pet prstiju sa malo izmenjenim brojem falangi (sl. 546).
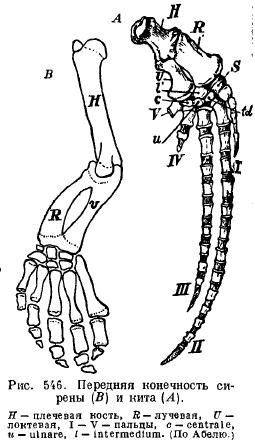
Zadnji udovi modernih kitova i sirena su toliko smanjeni da su ostali samo ostaci karlice (sl. 547 i 548).
Još jedan tip prilagođavanja udova kretanju u mahuni vidimo kod peronožaca (Pinnipedia). Ovdje rep gotovo nestaje, a stražnje noge se pomiču unatrag, igrajući ulogu motoričkog organa. Tri porodice ovog reda predstavljaju, u smislu stepena razvoja adaptacije na kretanje u vodi, tri stadijuma (ne genetski). Kod morževa i uhastih tuljana (Otariidae) stražnji udovi se još uvijek mogu savijati prema naprijed ispod tijela, ali kod morževa je struktura udova primitivnija, kraći su i pokretljiviji. Kod tuljana (Phocidae), zadnje noge se više ne mogu savijati prema naprijed ispod tijela, ali prsti (peraje) i dalje imaju kandže.
Kod životinja koje se ukopaju, udovi su također podvrgnuti vrlo jakim modifikacijama: posebno su se jako promijenili udovi tobolčarske krtice (Notoryctes typhlops). Ima dva grebena na lopatici, kao i mnoge životinje koje se ukopavaju, što je zbog snažnog razvoja mišića triceps brachii; postoji ključna kost, kao i svi sisari koji se kopaju. Na humerusu se nalazi veoma jak greben, koji se račvasto grana i služi za pričvršćivanje jakih (neophodnih za kopanje) deltoidnih i prsnih mišića; unutrašnji kondil ramena je visoko razvijen zbog snažnog razvoja mišića pregibača ulnarnog zgloba (proc. olecranon), koji služi kao pričvrsna tačka za mišić tricepsa; zglob je posebno modificiran, postaje vrlo jak; 1. i 2. prst su pomjereni na dlan, a samo 3. i 4., opremljeni ogromnim kandžama, posebno 3., služe za kopanje; broj falangi prstiju je vrlo mali: 2 i 1. 5. metakarpal sa širokom ravnom kandžom, koja povećava površinu kopanja; u ravnini dlana nalazi se sesamoidna kost, za koju je pričvršćen mišić koji savija prste, koji igra važnu ulogu u kopanju; povećava snagu zgloba; kao i drugi oblici ukopavanja, simfiza pubisa je vrlo kratka; ischium se spaja sa poprečnim nastavcima oba zadnja sakralna pršljena; na butini, koljenu i tibiji nalaze se visoko razvijeni grebeni i izrasline - mjesta pričvršćivanja mišića; kosti tibije i fibule (tibia i fibula) su čvrsto spojene, kao kod zlatnog madeža (Chrysochloris); đon je okrenut prema van, kao kod Chrysochlorisa; 5. prst je savijen na plantarnu površinu, 5. nokat na stopalu je širok i lopatast, 4. je dugačak i srpast, 3. i 2. postaju sve kraći i širi; on grudne kosti visoko razvijena grebena, poput zlatne krtice, krtice (Spalax) i krtice (Talpa) (sl. 549).
Snažan razvoj grebena na kostima za pričvršćivanje mišića, skraćivanje karpusa, jaka veza karpalije, smanjenje nekih falangi i jačanje drugih koje imaju ulogu u kopanju - sve su ove osobine karakteristične za sisavce koji se kopaju uopšte. Ali ovisno o tlu u kojem životinja kopa, uočavat će se razlike. Prilikom kopanja u mekom tlu (na primjer, u krtici - Talpa) (Sl. 550), šaka se jako širi, djeluje kao lopata i ima tendenciju da razvije 6. prst. Prednji ud krtice je uporediv s lopatom, koja se koristi za kopanje u mekom tlu. Naprotiv, kod onih oblika koji kopaju po tvrdoj zemlji ruka postaje uža, u kopanju učestvuje samo nekoliko prstiju, uglavnom 3. i 4. koji se razvijaju jače od ostalih, kao kod Notoryctes i Chrysochloris. Ova četka je uporediva sa lopatom za kopanje u tvrdoj zemlji.
![]()
Istu stvar vidimo kod sisara koji kidaju čvrste strukture termita, na primjer, kod mravojeda (Myrmecophagus), u tamandua. Ove životinje hodaju, oslanjajući se na vanjsku ivicu stopala, sa uvučenim kandžama.
Adaptacije sisara na brzo kretanje na kopnu izazvale su vrlo značajne promjene na udovima. Kretanje po tlu, koje je bilo karakteristično za primitivne sisare, vršilo se oslanjanjem na cijelu ruku i stopalo; prvobitni oblici sisara bili su prvenstveno plantigradni ili plantigradni, odnosno oslanjali su se na cijelo stopalo prilikom hodanja, poput medvjeda. Ali takva struktura dopušta samo relativno sporo hodanje u poređenju sa antilopama ili konjima brzim nogama, ili zečevima, kengurima i jerboasima koji skaču. Tokom filogenetskog razvoja vidimo postepeno podizanje karpusa i tarzusa (karpusa i tarzusa) iznad zemlje, a zatim metakarpusa i metatarzusa, ali su falange i dalje u potpunosti dodirivale tlo. Na kraju su se falange ispravile, prednji i stražnji udovi počeli su se oslanjati na krajeve prstiju. Sada možemo pratiti faze ove evolucije korak po korak ne samo kod kopitara, već i kod predatorskih sisara koji su stekli sposobnost brzog trčanja. Kako se udovi ispravljaju, vanjski prsti više ne dodiruju tlo, prestaju funkcionirati i atrofiraju. To rezultira manjom površinom kontakta sa tlom. Promjene u distalnom dijelu udova dovele su do promjena u proksimalnom: karpalija i tarsalija se nalaze sve bliže jedna uz drugu, ulna i fibula su smanjene, udovi počinju da se kreću samo u sagitalnoj ravni, srednja metapodija se spajaju u jednu kost.
Dva razvojna puta vode do formiranja udova modernih kopitara. U jednom redu, osovina uda prolazi sredinom trećeg prsta i zadnji se najjače razvija, dok ostali postaju rudimentarni. To su kopitari (Perissodactyla), mezaksijalci (Mesaxonia). Kod drugih, osovina prolazi između trećeg i četvrtog prsta, koji se ravnomjerno razvijaju: artiodaktila (Artiodactyla), paraksijalna (Paraxonia). Bočni prsti su smanjeni. Razvoj kopitara je najjasnije vidljiv u filogenetskom razvoju udova konja (Sl. 551). Razvoju artiodaktilnog stanja doprinijelo je, između ostalog, i činjenica da mu je prethodila redukcija prvog prsta, tako da pri prelasku sa plantigradnog hoda gotovo da nije učestvovao, dok kod predaka jednopapka kod životinja razvoj petog prsta je u početku bio normalan,
Zanimljivo je da su i artiodaktili i kopitari nastajali u više navrata u evolucijskom procesu i u nekim slučajevima doveli do vrlo specijaliziranog tipa, na primjer, kod južnoameričkih Proterotlieriidae (izumrli).
Preostali prsti su prekriveni kopitima, koji se mogu dodatno razlikovati ovisno o načinu života i staništu. Dakle, koze koje ostaju na stijenama imaju strma i uska kopita i opremljene su oštrim rubom; kod sobova su vrlo dugačke, široko račvaste i služe kao svojevrsne skije koje sprečavaju životinju da padne u snijeg i močvare.
Istovremeno sa prstima, sa razvojem hodanja na prstima, dolazi do promjene na zglobu (carpus) i tarsusu (tarsus). Različito se javlja kod artiodaktila i kopitara zbog različite distribucije gravitacije. Konačno, u vezi s prelaskom na hodanje u luku, važnost lakatne kosti (ulna) kao oslonca za rame postepeno se smanjuje: ona se smanjuje ili spaja sa radijusom (radijusom). Na isti način, fibula se smanjuje i konačno nestaje u zadnjim udovima.
Značenje i priroda opisanih promjena jasno je vidljiva iz poređenja udova kamile i slona (Sl. 552). Kod slona šaka (stopalo) ostaje kratka, a rame i podlaktica (bedro i potkolenica) su produženi. Naprotiv, kod kamile se spojene metapodije produžuju; nadlaktica i podlaktica (kao i bedro i potkolenica) ostaju kratke. Zbog gubitka određenog broja kostiju, skelet ekstremiteta je pojednostavljen, potonji postaje lakši. Smanjenje prstiju dovodi do manjeg trenja, smanjujući rizik od oštećenja i rupture ligamenta. Fuzija metapodija stvara veću snagu. Ud slona je mnogo teži. Obilje kostiju u ruci zahtijeva spajanje svih prstiju kako bi se postigla stabilnost. opšta koža. Ud slona je stub velike snage, ali mu nedostaje lakoća udova kamile.
Drugi način da se brzo krećete po zemlji je skakanje. Ova se prilagodba više puta javljala u povijesti sisara i, štoviše, u širokom rasponu filogenetskih stabala: među tobolčarima - u klokanima (Macropus), među insektivorima - u skakačima (Macroscelidae) i, konačno, u različitim serijama razvoja glodavaca : porodica. zečevi (Leporidae) iz Duplicidentata; među vrstama nalik na dikobraz (Hystricomorplia) razvila se grupa dugonogih životinja (Pedetinae); među mišolikim vrstama (Myomorpha) imamo australskog jerboa (Conilurus), jerboa starog svijeta (Jaculus, Alactaga), američkih skakača (Zapus) i vrećastog skakača (Dipodomys).
Razvoj sposobnosti skakanja povezan je s produžavanjem stražnjeg ekstremiteta, odnosno metatarzalnih kostiju. Osim toga, dolazi do gubitka dodatnih vanjskih prstiju, a tarzalne kosti su međusobno vrlo čvrsto povezane. Primjeri su stopala zeca (Alactaga) i pravog jerboa (Dipus).
Prednje noge Alactage su male i kratke. Zec ih koristi prilikom kopanja, ali ne dodiruje zemlju sa njima kada skače. Skakanje se izvodi samo uz pomoć zadnje noge pa čak i bez pomoći repa. Potonji dodiruje tlo samo u mirnom položaju, kada se životinja oslanja na rep. Skelet noge ima posebnost da se metatarzalne kosti 2., 3. i 4. prsta spajaju u jednu kost. Krajevi metatarzalnih kostiju nisu spojeni ispod, a medijan (3. prst) strši dalje naprijed. Od falangi povezanih s njima, srednje su opet najduže. Prvi i peti prsti su zaostali i ne dodiruju tlo. Oba ova prsta imaju istu dužinu zbog činjenice da metatarzalna thumb duže od 5. Prvi ima 2, drugi - 3 falange.
U Dipusu je specijalizacija otišla još dalje: 1. prst je netragom nestao, a od 5. je ostao samo rudiment. Srednji prst dvostruko slabiji od ova dva, a tek nešto duži od njih. Prsti su paralelni. Srednji prst je smanjen jer njegovu funkciju preuzimaju 2. i 4. čiji krajevi pružaju potporu pri sjedenju i skakanju. U daljoj evoluciji, treći prst mora nestati: u Dipusu imamo nedovršeni tip adaptacije.
Rameni pojas sisari pojednostavljeno, sastoji se od velikih lopatice, koji imaju korakoidni izrast, i ključna kost(sl. 80, 81). U embrionalnoj fazi razvoja i kod odraslih monotremesa coracoid je nezavisna kost.
Spatula koji se nalazi na vrhu grudi, ima trokutasti oblik i visok izrast (greben) sa akromijalni proces vezivanja mišića. Na mjestu korakoidnog nastavka i akromijalnog nastavka lopatice nalazi se glenoidna jama za artikulaciju prednjeg uda.
Ključna kost u obliku štapa služi za spajanje ramena i prsne kosti. Ključna kost je najrazvijenija kod životinja koje izvode složene pokrete prednjim udovima (šišmiši, krtice, primati, glodavci itd.). Kod mnogih vrsta koje karakteriziraju monotoni pokreti (papkari, psi), ključna kost je odsutna. Općenito, pojas prednjih ekstremiteta povezan je sa aksijalnim skeletom preko mišića i ligamenata.
Zdjelični pojas (Sl. 82 A) sastoji se od uparenih bezimene kosti, formiran artikulacijom ilijačne, ischijalne i stidne kosti. karlica zatvorenog tipa a kroz ilijačne kosti je čvrsto povezan sa sakralni region kičma. Kada se elementi karlice srastu, to se događa pubična fuzija i formira se acetabulum za artikulaciju zadnjeg ekstremiteta. U donjem dijelu neimenovane kosti nalazi se obturator foramen.
Skelet parnih udova
Udovi sisara, kao i kod drugih kopnenih kralježnjaka, su petoprstog tipa i predstavljaju tročlanu polugu čiji su svi dijelovi međusobno pokretno povezani.
Prednji ud obuhvata rame, podlaktica i šaka(Sl. 81). Brahijalna kost dobro razvijena, ima zaobljenu glavu za pričvršćivanje za pojas ekstremiteta. Podlakticačine dvije kosti - radijalni i ulnarni, imajući olecranon na vrhu. Na spoju ramena i podlaktice se nalazi lakatnog zgloba.
Četka obuhvata tri odeljenja - zglob, metakarpus i falange. Ručni zglob sastoji se od 8-10 elemenata raspoređenih u tri reda . Pastern uključuje pet kostiju (prema broju prstiju). Svaki prst ima tri falange, osim prvog prsta koji se sastoji od dvije falange. Za razliku od interkarpalne artikulacije gmizavaca i ptica, pokretna joint kod sisara se nalazi između podlaktice i šake(kao vodozemci).
Zadnji ud sisari takođe imaju tri odeljenja - butina, potkolenica i stopalo(Sl. 82 B). Femur masivan i sa zaobljenom glavom umetnut u acetabulum karličnog pojasa. Tibija sadrži dvije kosti - tibije i fibule. Veća tibija čini unutrašnji dio potkolenice, a fibula čini njen vanjski dio.
Između butine i potkoljenice nalazi se joint, pokriveno kneecap, koje su evolucijski formirale njihove okoštale mišićne tetive. Površine zglobnih kostiju prekrivene su hrskavičnim formacijama. Zglob skočnog zgloba Kod sisara se nalazi između donjih krajeva tibije i proksimalnog dijela stopala.

Rice. 82. Karlični pojas (A) i zadnji ud (B) lisice:
1 – ilium, 2 – ishium, 3 – stidne kosti, 4 – acetabulum, 5 – femur, 6 - patela, 7 - tibija, 8 - fibula, 9 - tarsus, 10 - metatarsus, 11 - falange, 12 - talus, 13 - kalkaneus
Tarzalni elementi stopala su raspoređena u tri reda i formiraju izrasline - calcaneus, karakterističan za sisare. Metatarsus sastoji se od pet kostiju za koje su pričvršćene falange prstiju. Kao i u prednjim udovima, u svakom prstu ih ima po tri, osim prvog (unutrašnjeg), koji ima samo dvije falange.
Sisavce karakterizira poseban raspored udova u odnosu na tijelo: nalaze se ispod tijela, što doprinosi razvoju veće manevarske sposobnosti pokreta. Gde lakatnog zgloba usmjeren prema stražnjoj strani, a zglob koljena je, naprotiv, orijentiran prema naprijed.
Zbog raznolikosti vrsta sisara, njihov način života i načini kretanja značajno variraju relativna dužina udova, odnos sastavnih elemenata, konfiguracija kostiju, broj prstiju itd. Na primjer, kod slepih miševa dužina prednjeg uda, posebno falange prstiju, naglo je povećana, služeći za pričvršćivanje letne membrane.
Naprotiv, prednji ekstremitet mladeža je skraćen, palmarni dio šake je proširen, a šesti prst je dobro razvijen. Kod kitova je ruka oštro izdužena zbog povećanja broja falangi prstiju, zbog čega je ud dobio veliku sličnost s perajama ribe s režnjevima. Prsti životinja s velikom biomasom i brzim trčanjem (parnoprsti, parnoprsti) značajno su smanjeni i modificirani.
Vanjska struktura psa
Pas je bio prva domaća životinja. Čovek ga je ukrotio u davna vremena. Pas je pomogao primitivnom čoveku tokom lova, čuvala je njegov dom. Danas su poznati radni, lovački i ukrasni psi, kao i mješanci (mješanci). Psi pomažu u zaštiti granica naše domovine, i za vrijeme Velikog Otadžbinski rat tražio ranjenike, pomagao u izviđanju i komunikaciji. Naučnici provode istraživanja o vitalnim funkcijama tijela na psima. Pas pre čoveka posjećenog prostora.
Tijelo psa, poput pastira ili haskija, je vitko i mišićavo. Njegovi udovi nisu smješteni sa strane tijela, kao kod reptila, već ispod tijela, tako da tijelo životinje ne dodiruje tlo. Pas hoda, oslanjajući se na prste sa jakim kandžama. Fleksibilan vrat omogućava veću pokretljivost glave. Usta životinje ograničena su pokretnim usnama - gornjom i donjom. Iznad gornja usna postoji nos sa par vanjskih nosnih otvora - nozdrva. Oči imaju dobro razvijene kapke. Mikajuća membrana (treći kapak) kod psa je, kao i kod svih sisara, nedovoljno razvijena. Od svih životinja imaju samo sisari vanjskog uha - Ušna školjka. Vanjsko uho psa je veliko i pokretno.
Pokrivači sisara
Koža sisara je jaka i elastična, u većini životinja u njoj se nalaze osnove dlake koje čine karakterističnu osobinu ove klase kralježnjaka. linija kose . Postoje debeli i duga kosa — awn i one kraće, mekše - underfur, ili poddlaka. Gruba i jaka osa štiti poddlaku i kožu od oštećenja. Poddlaka, koja zadržava mnogo vazduha, dobro zadržava telesnu toplotu. Osim kičme i podloge, životinje razvijaju veliku dlaku - organe dodira.
Dlaka sisara, poput perja ptica i krljušti reptila, sastoji se od rožnate supstance. Pas se, kao i mnogi drugi sisari, dva puta godišnje linja - dio dlake mu opada i zamjenjuje se novom. Baza dlake nalazi se unutar posebnog folikula dlake u koji se otvaraju kanali obližnjih dlačica. lojne žlezde. Njihovi sekreti podmazuju kožu i kosu, čineći ih elastičnima i ne navlaženim vodom.
Slika: Građa kože i dlake sisara
U koži većine životinja nalaze se i znojne žlezde . Znoj isparava sa površine tela, hladeći ga. Zajedno sa znojem, višak soli i ureje se također uklanja iz tijela; tako znojne žlezde igraju ulogu dodatnih organa za izlučivanje.
Pas ima malo znojnih žlezda i hlađenje tela se postiže pojačanim disanjem.
Na krajevima prstiju sisara nalaze se rožnate kandže, nokti ili kopita. Ponekad se na glavi razvijaju i rožnate formacije (rogovi na nosorogama, antilopama, goveda itd.) ili na repu (na primjer, rožnate ljuske kod pacova).
Skelet sisara
Skelet sisara sastoji se od istih dijelova kao i kostur drugih kičmenjaka. Lobanju životinja odlikuje se većom lobanjom, koja je povezana s velike veličine mozak. Vrlo je tipično za sisare da imaju 7 vratnih pršljenova. I žirafe dugog vrata i kitovi imaju isti broj vratnih pršljenova. Torakalni pršljenovi(obično ih ima 12-15) zajedno sa rebrima i prsnom kosti čine jaku prsa. Masivni pršljenovi lumbalnog regiona su pokretno zglobljeni jedan s drugim. U ovom dijelu torzo se može savijati i savijati. Broj lumbalnih pršljenova varira između različite vrste(2-9), pas ih ima 6. Sakralni dio kičma (3-4 pršljena) se spaja sa karličnim kostima. Broj kralježaka u kaudalnom području (od tri do nekoliko desetina) ovisi o dužini repa.

Slika: Skelet sisara na primjeru psa
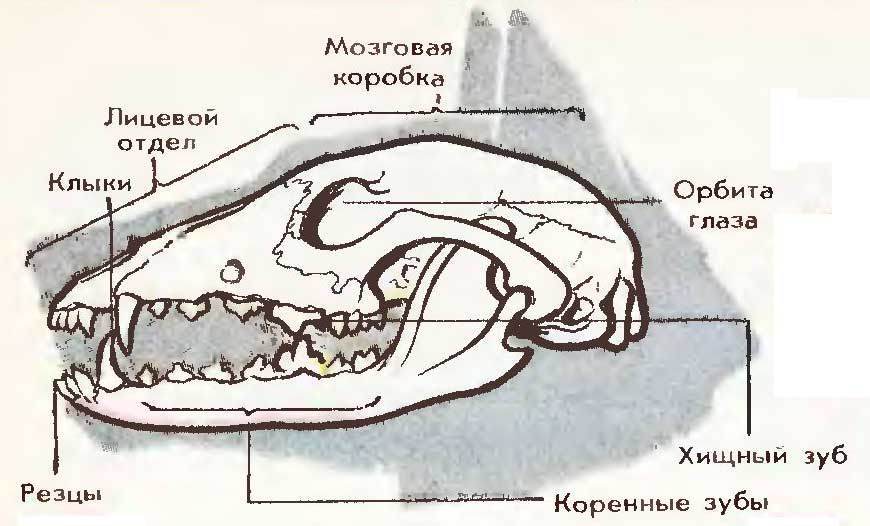
Uzorak: pseća lobanja
Pojas prednjih nogu sisara sastoji se od dvije lopatice na kojima su pričvršćene vranske kosti i dvije ključne kosti. Ključne kosti psa nisu razvijene. Pojas za zadnje udove- karlica - formirana od tri para karlične kosti. Skeleti udova različitih vrsta sisara i gmizavaca su slični, ali su detalji njihove strukture različiti kod različitih vrsta i ovise o životnim uvjetima životinje.
Mišići sisara
Većina sisara, uključujući pse, ima posebno razvijene mišiće na leđima, udovima i njihovim pojasevima. Pas može trčati velikim skokovima, savijati i ispravljati tijelo, naizmjenično se odričući od tla prednjim i zadnjim nogama. Jaki mišići pokrenuti donja vilica. Stežući čeljusti naoružani zubima, pas čvrsto drži plijen.

Slika: Mišići psa pokreću donju vilicu
Kičma se sastoji od vratnog, torakalnog, lumbalnog, sakralnog i kaudalnog dijela. Njegovo karakteristična karakteristika- platicelijalni (sa ravnim površinama) oblik pršljenova, između kojih se nalaze hrskavičasti intervertebralnih diskova. Gornji lukovi su dobro definisani. U vratnoj regiji ima sedam pršljenova, čija dužina određuje dužinu vrata; samo lamantin i lenjivac - Choloepus hof-fmani ih ima 6, a lenjivac - Bradypus 8-10. Cervikalni pršljenovi vrlo duga kod žirafa i vrlo kratka kod kitova, koji nemaju cervikalni presret. Do pršljenova torakalni rebra su zglobljena da formiraju grudni koš. Grudna kost koja ga zatvara je ravna i samo kod slepih miševa i vrsta koje se ukopavaju sa snažnim prednjim udovima (na primjer, krtica) ima mali greben (kobilicu), koji služi kao pričvrsna tačka prsnih mišića. U torakalnom dijelu ima 9-24 (obično 12-15) pršljenova, posljednjih 2-5 torakalnih pršljenova nose lažna rebra koja ne dopiru do grudne kosti.U lumbalnoj regiji ima od 2 do 9 pršljenova; Rudimentarna rebra se spajaju sa svojim velikim poprečnim nastavcima. Sakralni dio čine 4-10 spojenih pršljenova, od kojih su samo prva dva istinski sakralna, a ostali kaudalni. Broj slobodnih kaudalnih pršljenova kreće se od 3 (kod gibona) do 49 u dugorepog guštera.
Stepen pokretljivosti pojedinih pršljenova varira. Kod malih životinja koje trče i penjaju velika je, po cijeloj dužini kralježnice, pa se njihovo tijelo može savijati u različitim smjerovima, pa čak i sklupčati u klupko. Pršljenovi torakalnog i lumbalne regije kod velikih životinja koje se brzo kreću. Kod sisara koji se kreću na zadnjim nogama (klokani, jerboasi, skakači) najveći pršljenovi nalaze se u podnožju repa i sakruma, a dalje prema naprijed njihova veličina se sukcesivno smanjuje. Kod kopitara su, naprotiv, pršljenovi, a posebno njihovi spinovi nastavci, veći u prednjem dijelu torakalnog dijela, gdje su za njih pričvršćeni snažni mišići vrata i dijelom prednjih udova (
Slični članci
-
Kako je unutra uređena pravoslavna crkva?
Gdje su se molili prvi kršćani? Šta su oktogon, transept i naos? Kako je strukturiran hram u šatorima i zašto je ovaj oblik bio toliko popularan u Rusiji? Gdje se nalazi najviše mjesto u hramu i o čemu će vam freske govoriti? Koji se predmeti nalaze u oltaru? Hajde da podijelimo...
-
Prepodobni Gerasim Vologdski
Glavni izvor biografskih podataka o monahu Gerasimu je „Priča o čudima Gerasima Vologdskog“, koju je napisao izvesni Toma oko 1666. godine sa blagoslovom arhiepiskopa Vologdskog i Velikog Perma Markela. Prema priči...
-
Sveta ravnoapostolna Nina, prosvetiteljka Gruzije Mošti svete Nine
U jesen 2016. godine sestre Stavropigičkog manastira Svete Trojice Stefano-Mahrišči hodočastile su po svetim mestima Gruzije. Uoči proslave uspomene na svetog prosvetitelja Iverskog, nudimo vam foto reportažu o...
-
Sudbina ljudi rođenih 8. aprila
Ljudi rođeni na ovaj dan su izuzetno aktivni. Na život gledate kao na niz izazova i sve ih namjeravate riješiti. Ostvarujući svoje kreativne sposobnosti ili nastupajući kao šef velike korporacije,...
-
Nastavni čas "Poklonimo se tim velikim godinama" Scenario nastavnog časa za 9. maj
Pripremio nastavnik osnovne škole u MKOU Srednjoj školi br. Izberbash Nastavni sat. Cilj: Stvaranje neophodnih uslova za vaspitanje patriotskih osećanja kod mlađih školaraca, formiranje sopstvenog građanskog i patriotskog...
-
Formiranje kognitivnih vještina u osnovnoj školi
Govor Gusarove S.A. na sastanku nastavnika na temu: Formiranje kognitivnih veština učenja na časovima osnovne škole „Dete ne želi da uzima gotova znanja i izbegavaće onoga ko mu ga na silu zabija u glavu. Ali on svojevoljno...