De ce depinde scheletul membrelor pereche ale unui mamifer? Structura externă a mamiferelor, scheletului și mușchilor folosind exemplul unui câine domestic
Una dintre caracteristicile scheletului mamiferelor este mare dezvoltare cartilajul la animalele tinere și conservarea lui pe termen lung în perioada postembrionară de dezvoltare. Acest lucru se explică prin caracteristicile creșterii cartilajului și osului în legătură cu creșterea animalului însuși. Este cartilajul care crește cu întreaga sa masă (interstițial), creșterea are loc rapid și proporțiile părților pot fi menținute, în timp ce osul crește mai lent și doar de la suprafață, astfel încât părțile de schelet care apar să rămână mereu. la aceeași distanță și proporțiile părților se păstrează în timpul creșterii osoase imposibil. Acesta este motivul pentru care animalele tinere conțin întotdeauna mult cartilaj, și mai ales în locurile în care creșterea este intensă. Astfel, oasele lungi ale membrelor constau întotdeauna dintr-o parte mijlocie - diafiza, care se osifică mai întâi și capete care rămân cartilaginoase mult timp - epifiza. Osificarea diafizei începe în primul rând de la periferie - pericondral, adică datorită celulelor provenite din țesut conjunctiv, acoperind cartilajul, - din pericondriu; apoi se alătură prin osificare internă a cartilajului – osificare endocondrală; abia mai târziu, când apar carii în diafiză din cauza rupturii osoase măduvă osoasă, epifizele încep să se osifice endocondral; între diafize şi epifize rămâne însă mult timp cartilajul, cu ajutorul căruia osul creşte în lungime menţinând proporţiile (Fig. 530).

Coloana vertebrală mamiferele se caracterizează, spre deosebire de Sauropsida, prin absența articulațiilor libere între vertebre, cu excepția primei și a doua vertebre. Discurile de țesut conjunctiv elastic intervertebral se află între corpurile vertebrale. În general, acest lucru are ca rezultat mai puțină flexibilitate a coloanei vertebrale, ceea ce se datorează mișcării mai specializate a mamiferelor cu ajutorul membrelor.
Osificarea corpurilor vertebrale are loc prin epifizele cartilaginoase. Corpurile au fie suprafețe plane, fie ușor convexe, iar la majoritatea ungulatelor moderne sunt ușor opistocele. Bine dezvoltat pe vertebre, în special în zona pieptului, apofize spinoase, precum și apofize transversale care stau pe arcade; la baza acestuia din urmă se află procese articulare.

Numărul vertebrelor cervicale este de obicei 7 (Fig. 531), indiferent de lungimea gâtului, astfel încât lungimea gâtului, spre deosebire de păsări, depinde numai de lungimea vertebrelor. Doar lamantinul (Manatus) și o specie de lene (Choloepus hoffmani) au 6 vertebre cervicale, în timp ce lenesii cu trei degete (Bradypus) au 9. Aproape dispăruți din cauza stilului de viață acvatic, gâtul cetaceelor conține încă 7 plate, parțial topite. vertebre, în timp ce în gâtul unei girafe aceleași 7 vertebre sunt foarte lungi. Primele 2 vertebre cervicale sunt modificate în atlas și epistrofie, conectate prin articulații reale și în monotreme procesul odontoidîncă funcționează sub forma unui os special și nu este fuzionat, ca la alte mamifere, cu corpul epistrofeului. Coastele rudimentare ale regiunii cervicale fuzionează cu vertebrele, rezultând formarea unor procese transversale perforate. Fuziunea parțială sau completă a vertebrelor cervicale are loc la cetacee; la jerboa toate vertebrele cervicale fuzionează, cu excepția celei de-a 7-a, în alunița marsupial (Notoryctes) a 2-a la a 6-a vertebre, în aluniță - de la a 3-a la a 4-a.
Numărul de vertebre toracice, care se caracterizează prin atașarea coastelor de ele, variază de la 9 [la un cetacee (Hyperoodon) și armadillo (Tatusia)] la 25 [la un leneș], dar de obicei sunt 12-13 dintre ele . Vertebrele toracice au procese bine dezvoltate: spinoase (proc. spinosus), transversale sau diapofize (proc. transversus), articulare sau zigapofize (proc. articularis); Coastele sunt articulate de capetele lor cu corpul uneia sau două vertebre adiacente și de tuberculii lor la procesele transversale. Sunt formate din două părți: o coastă osoasă - proximală și o coastă cartilaginoasă - distală. Doar coastele vertebrelor toracice anterioare ajung la stern, coastele posterioare se termină liber ( coaste false). Vertebrele lombare, care se caracterizează prin absența coastelor care sunt incluse aici în procesele transversale, la mamifere variază de la 2 la 9, de obicei sunt 6-7.
Vertebrele sacrale ale mamiferelor sunt acele vertebre cu care, fuzionate, se articulează ilium. Există 1-10 astfel de vertebre, dar doar 1-2 dintre ele sunt vertebre sacrale adevărate, adică acelea în care articularea cu pelvisul are loc prin osificări independente corespunzătoare coastelor sacrale; vertebrele sacrale rămase sunt false, adică vertebrele caudale fuzionate cu sacrul.
Numărul vertebrelor incluse în regiunea caudală variază enorm: de la 3 (lilieci, giboni) la 49 [șopârlă (Manis macrura)].
Sternul(sternul) mamiferelor se dezvoltă din fuziunea capetelor ventrale ale coastelor, care formează apoi o singură placă nepereche, formată dintr-un număr de secțiuni (Fig. 532).

Conectat la clavicule (dacă există) și la prima pereche de coaste, extins sectiunea anterioara sternul se numește manubrium (praesternum, manubrium); Urmează o secțiune numită corp (mezostern, corpus), deseori descompusă într-o serie de oase, situate succesiv între perechi succesive de coaste; chiar mai în spate se află așa-zisa procesul xifoid(xiphisternum). Nu toate coastele sunt legate de stern, un număr mai mult sau mai puțin semnificativ de coaste false se termină liber (Fig. 533).

La mamiferele cloacale (Monotremata), datorită prezenței coracoizilor, care sunt absenți la toate celelalte mamifere, cartilajul presternal (prosternul) este așezat în fața manubriumului sternului, care este în legătură cu coracoizii, care este apoi înlocuit de osul în curs de dezvoltare - episternul, care aduce mamiferele cloacale mai aproape de reptile.
Scull Mamiferele au o serie de caracteristici care diferă de craniile reptilelor. Aceste trăsături nu au apărut însă toate deodată, ci prin schimbări treptate, care pot fi observate studiind craniile șopârlelor fosile (Theromorpha), în special din ordinul Theriodontia. Craniul de mamifer este un craniu tropibazal, dar cu o porțiune interorbitală scurtată, făcându-l să arate ca unul platibazal, cu un craniu mare, cu o tendință clară ca oasele individuale să se contopească,
Dispunerea oaselor în craniul mamiferelor poate fi apreciată din diagrama atașată (Fig. 534).
Pe oasele occipitale laterale, spre exterior de tuberculii articulari, există de obicei un proces occipital (proc. paroccipitalis), mai ales bine dezvoltat la rozătoare și multe ungulate. În fața tuberculilor articulari există de obicei o deschidere pentru ieșirea nervului hipoglos (XII) (pentru. condiloideum anterius).
În cartilajul capsulei auditive sunt așezate mai multe osificații, care însă foarte devreme se contopesc în două oase: cel situat în față - petrosum - și cel situat în exterior și în spate - mastoidul (mastoideum); ambele aceste oase, fuzionate, dau naștere petromastoidului, sau pur și simplu osului petros (petromastoideum, petrosum sau perioticum), care înconjoară labirintul auditiv intern. Perioticum se mărginește în spate cu principal și lateral oasele occipitale, iar în partea anterioară cu oasele sfenoid și pterigosfenoid; întrucât vasele și nervii trec între aceste oase și osul petros, aici rămân goluri semnificative: în spate - foramenul zdrențuit posterior (pentru. lacerum posterius), sau foramenul jugular (pentru. jugulare) pentru trecerea glosofaringianului (IX). ), ramuri ale nervului vag (X) și pentru nervul accesoriu (XI), precum și pentru cel intern vena jugulară, iar în față - gaura zdrențuită anterioară (pentru. lacerum anterius), prin care internă artera carotida. Uneori, legătura dintre osul petros și craniu este atât de slabă încât poate fi ușor separată împreună cu osul atașat de acesta. osul timpanic sau chiar în timpul macerării craniului acesta cade complet, așa cum este cazul cetaceelor.
În afara regiunii urechii, se formează două oase tegumentare. Unul dintre ele, a cărui origine încă nu este suficient de clarificată, se dezvoltă în domeniul extern canalul urechii sub forma unui os mic semicircular. Acesta este osul timpanic (timpanicum), care este probabil un omolog al osului unghiular (angulare) al maxilarului inferior al reptilelor, care s-a mutat în regiunea parotidă împreună cu alte oase ale maxilarului inferior din cauza modificărilor care au avut loc în maxilar. articulare. Situat inițial sub forma unui os semicircular sau inelar în zona canalului auditiv, deseori crește și formează, de obicei împreună cu osul petros și uneori cu osul solzos, canalul auditiv extern (meatus auditorius externus). Există tensiune pe osul timpanic timpan. Mai adânc în interior, osul timpanic poate participa și la formarea pereților urechii medii. Adesea, osul timpanic de pe partea inferioară a regiunii auditive se umflă într-o bulă cu pereți subțiri (bulla ossea).
Al doilea os de origine dermică care participă la formarea scheletului regiunii urechii este osul solzos (squamosum); a primit o dezvoltare și o semnificație excepționale la mamifere datorită atașării maxilarului inferior de acesta. La maimuțe și oameni, toate aceste oase ale regiunii urechii sunt fuzionate împreună pentru a forma osul temporal complex (temporal); Osul solzoasă constituie aici doar o parte, și anume solzii (squama temporalis) osul temporal, în timp ce osul timpanic este partea timpanică (pars tympanicum), iar parotida propriu-zisă oase cartilaginoase formează părțile pietroase (pars petrosa) și mastoid (pars mastoidea). Gradul de dezvoltare a osului scuamos și participarea acestuia la formarea pereților carcasei cerebrale și ai canalului auditiv sunt foarte diferite, dar osul scuamos are întotdeauna o suprafață articulară (cavitas glenoidea) pentru articularea maxilarului inferior. Astfel, la mamifere, spre deosebire de toate celelalte vertebrate, există o articulație originală foarte specială a maxilarului inferior: nu la osul pătrat, ci la osul scuamos al craniului. Suprafața articulară pentru maxilarul inferior se află pe baza procesului zigomatic (proc. zygomaticus) extinzându-se spre exterior și înainte de la solzi. În monotreme, deasupra suprafeței articulare, este străpuns de canalul temporal (can. temporalis), exprimat mai ales bine la ornitorinc. În spatele fosei articulare există un proces (proc. postglenoideus).
Anterior regiunii auditive, baza craniului este formată din principal osul sfenoid(basisphenoidale), pe a cărei suprafață interioară există o depresiune mai mult sau mai puțin semnificativă - așa-numita sella turcică (sella turtica), în care se află glanda pituitară. Pe părțile laterale ale osului sfenoid principal se află, formând părțile inferioare ale peretelui anterior al craniului, oasele pterigosfenoide (alisphenoidea), de obicei fuzionate cu acesta într-un singur os, formând aripi mari (alae magnae) ale acestui os. Oasele pterigosfenoide sunt dotate cu două deschideri: oval (pentru. ovale) pentru trecerea celei de-a treia ramuri nervul trigemen(V3) și rotund (pentru. rotundum) pentru trecerea celei de-a doua ramuri a aceluiași nerv (V2). Cu toate acestea, primul se contopește adesea cu foramenul zdrențuit anterior (for. lacerum anteris), iar al doilea se contopește adesea cu foramenul sfenorbital (for. sphenorbitale), delimitat simultan atât de osul pterigosfenoid, cât și de osul orbital-sfenoid aflat în fața acestuia. aceasta. Prin această deschidere trec oculomotor (oculomotorius III), trohlear (trohlearis IV), abducens (VI) și prima ramură a nervului trigemen.
Calota craniană aici este acoperită de oasele parietale (parietale), care adesea cresc împreună (în monotreme, unele marsupiale și ungulate); între ele și osul occipital superior mai sunt așezate două oase, contopindu-se într-un singur os interparietal (interparietale), care în unele ordine se păstrează ca os independent (cetacee, marsupiale, hyraxs, aardvarks, aripi lânoase și unele rozătoare), în timp ce la altele se contopește sau cu parietalul (la unele rozătoare, sireniene și rumegătoare), sau cu osul occipital superior (la carnivore, animale unicopilate, primate).
În fața osului sfenoid principal se află osul sfenoid anterior (praesphenoideum) cu o depresiune pentru chiasma nervilor oftalmici; pe părțile laterale ale acestuia, de obicei fuzionate cu acesta, se află oasele orbital-sfenoide (orbitosphenoidea) cu o deschidere pentru trecerea nervilor orbitali (for. opticum), numite aripi mici (alae parvae) în cazul fuziunii cu oasele sfenoide anterioare și principale. După cum am menționat mai sus, între ambele oase pterigoide de fiecare parte există deschideri sub formă de fante prin care trec nervii muschii ochilor iar prima ramură a nervului trigemen (pentru. sphenorbitale).
Peste acest rând de oase corespund oasele frontale tegumentare (frontale) - de obicei oase pereche bine dezvoltate, uneori fuzionate și purtătoare de tije la bovidi și cioturi pentru coarne la căprioare. Acoperind craniul, oasele frontale sunt limitate deasupra și cu interior orbite, formând adesea procese: preorbitale (proc. praeorbitalis și postorbitale (proc. postorbitalis). Uneori în jurul ochilor se formează un inel periorbital datorită fuziunii osului zigomatic (jugale) cu osul frontal (unele insectivore și carnivore, rumegătoare). , animale cu o singură copită, prosimieni).La maimuțe, procesul de separare a alveolei orbitale de alveola temporală merge atât de departe încât se formează un perete osos între orbită și cavitatea temporală.
În peretele anterior al orbitei se află un mic osicul lacrimal tegumentar (lacrymale), perforat de deschiderea canalului lacrimal (pentru. lacrymale).
În regiunea nazală, în capsula nazală cartilaginoasă se formează mai multe osificări. În primul rând, de-a lungul septului nazal cartilaginos, în mijloc se dezvoltă un sept osos (lamina perpendicularis), corespunzător osului olfactiv mediu (mesetmoideum); în al doilea rând, în zidul din spate capsula nazală și în peretele anterior al cavității cerebrale se dezvoltă două plăci laterale, perforate de numeroase [doar la ornitorinc aceste orificii sunt înlocuite cu una (pentru. olfactorium)] deschideri pentru trecerea nervului olfactiv - plăci cribriforme (lamina). cribrosa), corespunzătoare oaselor etmoidale laterale (ethmoidalia lateralia) . Din septul nazal, osificarea se extinde din partea dorsală de-a lungul laturilor, trece de-a lungul pereților laterali ai capsulelor nazale și se termină în partea inferioară, formând numeroase proeminențe din lateral în cavitatea nazală. Acestea sunt conchele olfactive (etmoturbinalia), care împreună cu plăcile laterale și cavitățile structurii celulare formează uneori un labirint nazal foarte complex. Poziția cochiliilor poate fi văzută din desenul schematic atașat (Fig. 535). Din osul nazal tegumentar (nasale), care acoperă această zonă deasupra, cochiliile (nasoturbmalia) atârnă în cavitatea nazală, de asemenea, câte una pe fiecare parte. De jos încolo sept nazal se suprapune unui os nepereche, considerat anterior a fi un vomer. Cu toate acestea, studii recente au arătat că, pe lângă acest os tegumentar nepereche, la baza craniului se dezvoltă două oase pereche, numite prevomer; și întrucât la reptile vomerul este prezent sub forma unui os pereche, iar osul nepereche din această zonă era parasfenoidul, atunci vomerul mamiferelor poate fi considerat omologul acestui ultim os la reptile, iar prevomerii pot fi considerați vomere. În fața pereților cavității nazale se află oasele maxilarului (maxilare), pe care se formează cochilii care cresc în cavitatea nazală; acestea sunt cochiliile maxilarului (maxiloturbinale).

Partea frontală a craniului se termină în față cu oasele premaxilare (praemaxillare). La maimuțele antropomorfe și la oameni, acest os fuzionează foarte devreme cu osul maxilarului, iar la chiroptere este redus la un os mic sau chiar absent cu totul. Cu procesul său vertical, osul premaxilar limitează deschiderea nazală în față și se extinde între osul nazal și osul maxilar în spate, uneori până la osul frontal. Mai jos, oasele premaxilare iau parte la formare palatul tare. Ele limitează deschiderile anterioare și laterale ale canalului palatin (Stenson) (pentru. incisiva) pe palatul dur, conectând cavitatea bucală cu orga lui Jacobson. Procesele oaselor premaxilare aflate în afara acestor deschideri palatonazale sunt suprapuse pentru fiecare os în mod independent, sunt probabil omologi ale vomerului pereche de reptile (praevomer) iar în ornitorinc se păstrează ca oase independente. Incisivii sunt localizați pe osul premaxilar.
Oasele maxilarului, așa cum sa menționat deja, limitează fundul cavității nazale, formând palatul dur cu procese orizontale. În interiorul cavității nazale, învelișul inferior (maxiloturbinae) crește până la pereții săi laterali, formați din oasele maxilarului, și se formează ca un os independent.
Continuarea spatelui palat dur sunt procesele orizontale ale oaselor palatine (palatin), care limitează pasajul nazal intern ca pereții laterali. Ele stau vertical și sunt, de asemenea, unite în spate prin oase pterigoidee vertical, care la om fuzionează cu osul sfenoid, formând procesele pterigoide ale acestuia (proc. pterygoidei).
Partea anterioară a craniului suferă modificări semnificative în legătură cu trecerea la un stil de viață acvatic la cetacee, sirenieni și în legătură cu dezvoltarea trunchiului la elefanți.

La cetacee (Fig. 536), nările sunt mutate mult înapoi în regiunea interorbitală și duc într-un canal vertical, care se deschide cu coaele în cavitatea bucală; oasele nazale se află în spatele nărilor, suprapunând oasele frontale și aproape adiacente osului interparietal; De asemenea, oasele maxilarului se suprapun pe oasele frontale, extinzându-se aproape până la oasele parietale, puternic deplasate în lateral de osul interparietal.
La sirene are loc aceeași mișcare în spatele nărilor, dar nu atât de departe; oasele nazale de aici sunt și mai reduse și pot chiar să lipsească (Fig. 537).

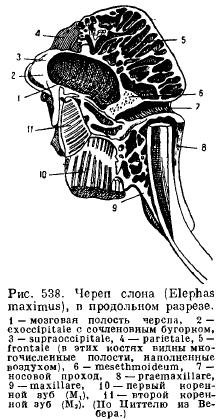
La elefanți, odată cu dezvoltarea excesivă a colților și a molarilor puternici și datorită prezenței unui trunchi, dispunerea normală a oaselor este mult modificată: nările sunt puternic împinse înapoi, și de aceea canalul nazal ia o poziție aproape perpendiculară; oasele frontale sunt foarte scurte, oasele premaxilare sunt foarte mari și se extind mult înapoi; oasele maxilarului sunt deplasate sub baza craniului; întregul craniu devine foarte masiv cu pneumaticitatea extrem de dezvoltată a oaselor (Fig. 538).
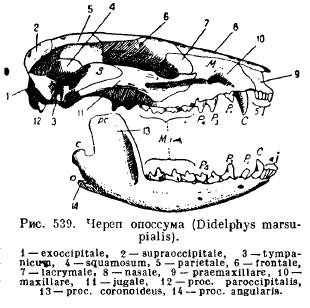
Maxilarul inferior al mamiferelor, spre deosebire de toate celelalte vertebrate, se dezvoltă pe fiecare parte dintr-un singur os dentar (dentale); ambele jumătăți fie rămân legate între ele prin cartilaj, sutură, fie cresc împreună (animale cu o copită, elefanți, lilieci, maimuțe). Fiecare jumătate este formată dintr-un corp orizontal (corpus), pe care se așează dinții și o parte ascendentă cu procese: partea anterioară. procesul coronoid(proc. coronoideus), un proces articular situat în spatele lui (proc. articularis) și la mamiferele inferioare un proces unghiular înapoi (proc. angularis); la majoritatea marsupialelor și la unele rozătoare, acest proces se transformă într-o placă care se extinde spre interior (Fig. 539).
Această structură a maxilarului inferior și atașarea sa particulară de craniu ne conduc la întrebarea originii acestor caracteristici la mamifere. Poveste dezvoltarea individuală iar datele paleontologice oferă un răspuns la această întrebare. La embrionii de mamifere se formează cartilajele palatoquadrate și Meckel. Cu toate acestea, osificarea cartilajului palatoquadrate are loc independent de articulația maxilarului inferior și nu duce la formarea unui os pătrat, ca la reptile, și va da naștere unui nou os auditiv - incusul (incusul), în timp ce Meckel. cartilajul, osificându-se în porțiunea sa posterioară, dă naștere unuia nearticular.os (articulare), ca și alți gnatostomi, iar al treilea os auditiv este malleus (malleus). Primul osicul auditiv, stapes (stapes), corespunde coloanei de amfibieni, reptile și păsări și, după cum știm deja, este o suspensie modificată (hyomandibulare), adică partea proximală a arcului hioid. La fel ca coloana (columella auris), etrierul acoperă fereastra ovala labirint auditiv, și este perforat, ca la unele reptile, cu o deschidere prin care trece artera facială (a. facialis) la unele mamifere inferioare (Fig. 540 și 541).

Astfel, la mamifere, osul pătrat se transforma într-un incus, osul articular într-un maleus; și întrucât un mic os tegumentar crește până la malleus, formând procesul său mijlociu, se crede că acesta din urmă corespunde osului articular tegumentar (goniale) al reptilelor. Osul unghiular tegumentar al reptilelor, situat adiacent acestui ultim os, trece și el în regiunea urechii, devenind osul timpanic (timpanicum). Aceste schimbari vizibil în diagrama atașată (Fig. 541).
Datele paleontologice confirmă posibilitatea transformărilor de mai sus. Printre șopârlele cu dinți de fiară (Theriodontia) se numără cele la care osul dentar primește o dezvoltare deosebit de mare, ajungând în spatele locului de articulare a maxilarului inferior cu manus pătrat. În același timp, scuamosul coboară de-a lungul pătratului până la aceeași articulație. Oasele dentare și scuamos se apropie astfel din ce în ce mai mult.
În cele din urmă, la unele mamifere, oasele dentare și scuamoase participă la articulație împreună cu oasele articulare și pătrate.
În ceea ce privește aparatul hioid al mamiferelor, acesta constă dintr-un corp, o parte bazală modificată (copula) a arcului hioid, coarne anterioare (cornua anteriora), formate din partea inferioară a arcului hioid și coarne posterioare (cornua posteriora) , transformat din primul arc branial. Coarnele anterioare constau din mai multe osificații (până la patru); segmentul orientat spre craniu crește adesea pe osul petros, formând un proces stiloid (proc. styloideus).
Al doilea și al treilea arc branhial au dat naștere formațiunii cartilajul tiroidian(cartilago thyreoidea.)
Centuri pentru membre. Centura scapulară a mamiferelor se caracterizează în primul rând prin reducerea oaselor corbului (procoracoideum, epicoracoideum, coracoideum), o tendință spre dispariția claviculei și o creastă bine dezvoltată a scapulei (spina scapulae), continuând în acromion.
Cu toate acestea, în monotreme, centura scapulară este foarte asemănătoare cu reptilele. Omoplatul este echipat cu o proeminență acromială, de care se alătură clavicula în față, iar oasele late și scurte de corb (coracoideum) care merg la stern sunt atașate la spate de scapula. În fața lor există procoracoizi largi (procoracoideum) sub formă de oase independente. Clavicula (claviculă) se conectează medial la epistern (Fig. 542). La alte mamifere, cracoizii se formează sub formă de două osificări, dar pierd legătura cu sternul și cresc până la scapula, formând un proces coracoid (proc. coracoideus); procoracoizii dispar complet (Fig. 543), claviculele dispar la unele carnivore și ungulate datorită adaptării la alergarea rapidă, iar la cetacee și sireniene datorită transformării membrelor anterioare în aripioare.


Brâul pelvin al mamiferelor are un aranjament tipic de elemente atunci când se formează; particularitatea sa este, în primul rând, prezența unui os suplimentar în acetabul (acetabulare), în al doilea rând, fuziunea tuturor acestor oase de fiecare parte într-un os fără nume (innominatum) și fuziunea oaselor pubiene și ischiatice, astfel încât un structură închisă se formează pe toate părțile există o gaură între aceste oase (pentru. obturatum) (Fig. 544). La monotreme și marsupiale, un os special este atașat de osul pubian de jos pentru a susține bursa (marsupiale).
Scheletul membrelorîn formele originale ale mamiferelor se datorează adaptării lor la mișcări rapide - alergarea cu ajutorul a patru membre, care sunt situate sub corp, astfel încât acesta din urmă să nu atârnă între membre, ca la majoritatea reptilelor, ci să se sprijine direct pe lor. Capacitatea de a alerga este legată de poziția diferită a membrelor la mamifere. Membrele lor sunt orientate astfel încât articulațiile cotului membrelor anterioare să fie îndreptate înapoi, iar articulațiile genunchilor membrelor posterioare să fie îndreptate înainte. La mișcare, membrele anterioare sunt aruncate înainte și trag corpul prin flexia articulațiilor încheieturii mâinii și ale cotului; Membrele posterioare împing corpul înainte prin îndreptarea piciorului la articulația genunchiului.

În ceea ce privește numărul de oase incluse în membre și locația acestora, scheletul membrelor formelor ancestrale de mamifere are o structură foarte tipică a unui membru primitiv cu cinci degete și a suferit modificări majore în cadrul clasei numai în acest proces. de adaptare la diverse tipuri de mişcări în conditii diferite mediu inconjurator.
Membrul anterior se caracterizează prin prezența în secțiunea distală humerus deschideri (pentru. entepicondilare), prezența unei proeminențe proximale ulna(olecranon), prezența unui os pisiform (pisiform) și fuziunea celui de-al patrulea și al cincilea oase carpien distal (carpalia distalia 4+6 = hamatum, sau uncinatum) și proximalul central cu intermediarul (dist. central + intermediu = lunatum sau semilunar). Datorită faptului că se folosește des nomenclatura oaselor încheieturii mâinii mamiferelor, adoptată în anatomia umană, vă prezentăm un tabel cu aceste denumiri într-o diagramă care indică locația oaselor.
Membrul posterior al mamiferelor se caracterizează în primul rând prin faptul că articulația genunchiului este rotită în față, coapsa are procese bine dezvoltate pe partea proximală - trohantere, inclusiv 2-3 (trohanter major, minor și tertius); în tibie tibiei mai dezvoltat, există os genunchieră(rotulă), protectoare articulatia genunchiului. Tarsul are doar două oase proximale, bine dezvoltate: talusul intern (talus = astragalus), format din fuziunea proximalului intermediar și central (intermediu + central proximal), și calcaneul extern (calcaneus), corespunzător tarsalului fibular. os (fibulare). Din osiculele rândului mijlociu, distalul central (centrale distale) s-a transformat într-un navicular (naviculare = scaphoideum). Osul tibial a fost fie redus, fie fuzionat cu acesta din urmă.
Aranjamentul oaselor tarsian și metatarsian și sinonimia numelor sunt următoarele:
În legătură cu adaptarea la mișcare în diverse medii, membrele mamiferelor au suferit modificări foarte semnificative, iar aceste modificări pot fi urmărite în mod deosebit clar pe schelet. Din păcate, nu avem date paleontologice despre structura membrelor strămoșilor mamiferelor. Nu există nicio îndoială că primele mamifere aveau membre slab specializate de tipul descris mai sus. Printre mamiferele noastre moderne, cele mai primitive marsupiale, de exemplu, opossums - șobolani marsupial (Didelphys) și insectivore care duc un stil de viață arboricol, au tocmai aceste membre. Cu toate acestea, ar fi greșit să concluzionăm din aceasta că strămoșii mamiferelor au fost cu siguranță locuitori ai pădurilor și, în plus, forme de cățărare în copaci. În plus, membrul opossum, deși este folosit pentru mișcarea de-a lungul ramurilor copacilor, nu este în niciun caz un membru arboricol specializat, tipic, dar își păstrează tipul universal cu cinci degete. Astfel de membre ar putea fi cel mai bine conservate la animalele care nu numai că alergau pe pământ, ci și se cățărău în copaci, precum și la animalele care urcau pe munți. Specializarea membrelor mamiferelor s-a produs concomitent cu diferențierea ordinelor și a altor unități sistematice mai mici și au avut loc convergențe.
Membrele suferă modificări deosebit de profunde atunci când se adaptează la zbor, înot în apă și se deplasează sub pământ; mișcările lente pe sol și un stil de viață arboricol nu se datorează specializării profunde.
La lilieci (Chiroptera), membrul anterior este transformat într-o aripă, iar membrana de zbor este întinsă între patru (2-b) degete foarte alungite, lipsite de gheare; primul deget este scurt și are o gheară puternic dezvoltată la capăt; acest deget nu joacă niciun rol în zbor, este folosit pentru cățărare; la încheietura mâinii, scafoidul, lunar și oasele centrale fuzionează într-unul singur; antebratul este alungit si este format dintr-o raza bine dezvoltata, in timp ce ulna este redusa si adera de raza. Membrana zburătoare acoperă și picioarele din spate. Între acestea din urmă, se dezvoltă diferit la lilieci (Fig. 545).
La mamiferele care înoată în apă, membrele se transformă în aripioare într-un mod similar, așa cum am văzut deja la reptile. Membrele balenelor și sirenelor s-au schimbat în mod deosebit dramatic. Spre deosebire de aripa, membrul anterior este scurtat semnificativ în partea sa proximală. La balene, oasele umărului și antebrațului au devenit plăci scurte și largi, oferind spațiu pentru atașarea mușchilor puternici. Toate secțiunile membrului sunt conectate nemișcat între ele, formând o vâslă puternică. În acest caz, numărul de falange poate fi mult crescut. Am văzut aceleași schimbări la reptile. Creșterea numărului de falange depinde de faptul că diafizele falangelor nu fuzionează cu epifizele și se osifică din centre separate. Această osificare separată se datorează procesului mai lent de osificare la aceste animale. Lentoarea procesului de osificare duce la faptul că majoritatea oaselor de la joncțiunile lor rămân cartilaginoase.
Înotătoarele sirenelor (Sirenia) sunt asemănătoare ca aspect cu cele ale cetaceelor: umărul și antebrațul sunt articulate astfel încât să se poată îndoi; există aproape toate oasele încheieturii mâinii și toate cele cinci degete cu un număr ușor modificat de falange (Fig. 546).
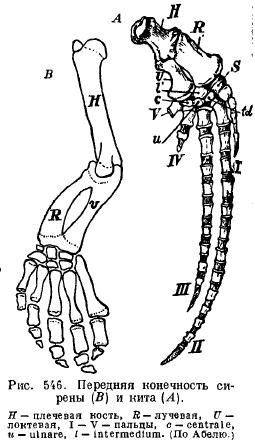
Membrele posterioare ale balenelor și sirenienilor moderni sunt atât de reduse încât rămân doar rămășițe din pelvis (Fig. 547 și 548).
Vedem un alt tip de adaptare a membrelor la mișcare în păstaie la pinipede (Pinnipedia). Aici coada aproape dispare, iar picioarele din spate se deplasează înapoi, jucând rolul unui organ motor. Cele trei familii din acest ordin reprezintă, din punct de vedere al gradului de dezvoltare de adaptare la mișcarea în apă, trei etape (nu genetic). La morse și focile urechi (Otariidae), membrele posterioare încă se pot apleca înainte sub corp, dar la morse structura membrelor este mai primitivă, sunt mai scurte și mai mobile. La foci (Phocidae), picioarele din spate nu se mai pot apleca înainte sub corp, dar degetele de la picioare (naboare) au încă gheare.
La animalele de vizuină, membrele suferă și ele modificări foarte puternice: membrele aluniței marsupiale (Notoryctes typhlops) s-au schimbat deosebit de puternic. Are două creste pe omoplat, la fel ca multe animale vizuitoare, ceea ce se datorează dezvoltării puternice a mușchiului triceps brahial; există o claviculă, la fel ca toate mamiferele vizuinătoare. Pe humerus există o creastă foarte puternică, ramificată în formă de furculiță și care servește la atașarea mușchilor deltoizi și pectorali puternici (necesarii săpăturii); condilul intern al umărului este foarte dezvoltat datorită dezvoltării puternice a mușchilor flexori ulnari (proc. olecranon), care servește ca punct de atașare pentru mușchiul triceps; încheietura mâinii este modificată în mod deosebit, devenind foarte puternică; Degetele 1 și 2 sunt mutate în partea palmară, iar la săpat se folosesc doar al 3-lea și al 4-lea, echipate cu gheare uriașe, în special al 3-lea; numărul de falange ale degetelor este foarte mic: 2 și 1. metacarpalul 5 cu gheara plată lată, care mărește suprafața de săpat; în planul palmei se află un os sesamoid, de care este atașat mușchiul care flectează degetele, care joacă un rol important în săpare; crește rezistența încheieturii mâinii; ca și alte forme de vizuină, simfiza pubisului este foarte scurtă; ischionul fuzionează cu procesele transversale ale ambelor ultime vertebre sacrale; pe coapsă, genunchi și tibie există creste și excrescențe foarte dezvoltate - locuri de atașare musculară; oasele tibiei și fibulei (tibia și peronéul) sunt ferm fuzionate, ca în alunița de aur (Chrysochloris); talpa este întoarsă spre exterior, ca la Chrysochloris; Al 5-lea deget de la picior este îndoit pe suprafața plantară, a 5-a unghie de pe picior este lată și în formă de picior, al 4-lea este lung și în formă de seceră, a 3-a și a 2-a devin mai scurte și mai late; pe sternul o creastă foarte dezvoltată, ca cea a aluniței de aur, a șobolanului aluniță (Spalax) și a cârtiței (Talpa) (Fig. 549).
Dezvoltarea puternică a crestelor pe oase pentru atașarea mușchilor, scurtarea carpului, conexiunea puternică a carpaliei, reducerea unor falange și întărirea altora care joacă un rol în săpare - toate aceste caracteristici sunt caracteristice mamiferelor îngropatoare în general. Dar in functie de solul in care sapa animalul se vor observa diferente. Când sapă în pământ moale (de exemplu, într-o aluniță - Talpa) (Fig. 550), mâna se extinde foarte mult, acționează ca o lopată și are tendința de a dezvolta al 6-lea deget. Membrele anterioare ale unei alunițe sunt comparabile cu o lopată, folosită pentru săpat în pământ moale. Dimpotrivă, la acele forme care sapă în pământ dur, mâna devine mai îngustă; doar câteva degete iau parte la săpătură, mai ales al 3-lea și al 4-lea, care se dezvoltă mai puternic decât altele, ca la Notoryctes și Chrysochloris. Această perie este comparabilă cu o cazma pentru săpat în pământ dur.
![]()
Același lucru îl vedem la mamifere care sfâșie structurile dure ale termitelor, de exemplu, la furnicar (Myrmecophagus), la tamandua. Aceste animale merg, sprijinindu-se pe marginea exterioară a picioarelor, cu ghearele înfipte.
Adaptările mamiferelor la mișcarea rapidă pe uscat au provocat modificări foarte semnificative la nivelul membrelor. Mișcarea pe sol, care era caracteristică mamiferelor primitive, era efectuată bazându-se pe întreaga mână și picior; formele originale de mamifere au fost în primul rând plantigrade sau plantigrade, adică se bazau pe întregul picior când mergeau, cum ar fi urșii. Dar o astfel de structură permite doar mersul relativ lent în comparație cu antilopele sau caii cu picior în picioare sau cu iepuri de câmp, canguri și jerboi. În timpul dezvoltării filogenetice, vedem o ridicare treptată a carpului și tarsului (carp și tars) deasupra solului, urmată de metacarp și metatars, dar falangele încă au atins solul în întregime. În cele din urmă, falangele s-au îndreptat, membrele anterioare și posterioare au început să se sprijine pe capetele degetelor. Putem urmări acum etapele acestei evoluții pas cu pas nu numai la ungulate, ci și la mamiferele prădătoare care au dobândit capacitatea de a alerga rapid. Pe măsură ce membrele se îndreaptă, degetele exterioare nu mai ating solul, încetează să funcționeze și suferă atrofie. Acest lucru are ca rezultat o suprafață de contact mai mică cu solul. Modificările în partea distală a membrelor au determinat modificări în cea proximală: carpalia și tarsalia sunt situate din ce în ce mai aproape una lângă alta, ulna și fibula sunt reduse, membrele încep să se miște numai în plan sagital, metapodiile mediane se îmbină într-un singur os.
Două căi de dezvoltare duc la formarea membrelor ungulatelor moderne. Într-un rând, axa membrului trece prin mijlocul celui de-al treilea deget, iar ultimul se dezvoltă cel mai puternic, în timp ce restul devin rudimentare. Acestea sunt ecvideele (Perissodactyla), mesaxiale (Mesaxonia). La altele, axa trece între al treilea și al patrulea deget, care se dezvoltă uniform: artiodactyla (Artiodactyla), paraxial (Paraxonia). Degetele laterale sunt reduse. Dezvoltarea ecvideelor este vizibilă cel mai clar în dezvoltarea filogenetică a membrelor calului (Fig. 551). Dezvoltarea stării de artiodactil s-a datorat, printre altele, faptului că a fost precedată de o reducere a primului deget de la picior, astfel că în timpul tranziției de la mersul plantigrad nu a luat parte aproape, în timp ce la strămoșii monocopițelor. animale, dezvoltarea celui de-al cincilea deget de la picior a fost inițial normală,
Este interesant că atât artiodactilele, cât și ecvideele au apărut în mod repetat în procesul evolutiv și, în unele cazuri, au condus la un tip foarte specializat, de exemplu, la Proterotlieriidae din America de Sud (disparută).
Degetele rămase sunt acoperite cu copite, care se pot diferenția și mai mult în funcție de stilul de viață și habitat. Astfel, caprele care stau pe stânci au copite abrupte și înguste și sunt dotate cu o margine ascuțită; la reni sunt foarte lungi, larg bifurcați și servesc ca un fel de schiuri care împiedică animalul să cadă în zăpadă și mlaștini.
Concomitent cu degetele, odată cu dezvoltarea mersului pe degete, apare o modificare la încheietura mâinii (carp) și tars (tars). Apare diferit la artiodactili și ecvidee datorită unei distribuții diferite a gravitației. În cele din urmă, în legătură cu trecerea la mersul cu arc, importanța ulnei (ulna) ca suport pentru umăr scade treptat: scade sau se contopește cu raza (raza). În același mod, peronéul este redus și în cele din urmă dispare în membrele posterioare.
Semnificația și natura modificărilor descrise sunt clar vizibile dintr-o comparație a membrelor unei cămile și ale unui elefant (Fig. 552). La un elefant, mâna (piciorul) rămâne scurtă, iar umărul și antebrațul (coapsa și piciorul inferior) sunt alungite. Dimpotrivă, la cămilă metapodiile topite se lungesc; brațul și antebrațul (precum și coapsa și piciorul inferior) rămân scurte. Din cauza pierderii unui număr de oase, scheletul membrului este simplificat, acesta din urmă devine mai ușor. Reducerea degetelor duce la mai puțină frecare, reducând riscul de deteriorare și ruptură a ligamentelor. Fuziunea metapodiilor creează o putere mai mare. Membrul elefantului este mult mai greu. Abundența oaselor din mână necesită conectarea tuturor degetelor pentru a obține stabilitate. piele generală. Membra unui elefant este o coloană de mare putere, dar lipsită de ușurința unui membru de cămilă.
Un alt mod de a te deplasa rapid pe sol este să sari. Această adaptare a apărut în repetate rânduri în istoria mamiferelor și, în plus, într-o mare varietate de trunchiuri filogenetice: printre marsupiale - la canguri (Macropus), printre insectivore - la săritori (Macroscelidae) și, în sfârșit, în diferite serii de dezvoltare a rozătoarelor. : familia. iepuri de câmp (Leporidae) din Duplicidentata; printre speciile asemănătoare porcului-sac (Hystricomorplia) s-a dezvoltat un grup de animale cu picioare lungi (Pedetinae); printre speciile asemănătoare șoarecelui (Myomorpha) avem jerboul australian (Conilurus), jerboul din Lumea Veche (Jaculus, Alactaga), săritorii americani (Zapus) și săritorii cu pungă (Dipodomys).
Dezvoltarea capacitatii de a sari este asociata cu alungirea membrului posterior, si anume oasele metatarsului. În plus, are loc pierderea degetelor exterioare suplimentare, iar oasele tarsale sunt legate între ele foarte ferm. Exemple sunt picioarele de iepure de câmp (Alactaga) și adevăratele jerboas (Dipus).
Picioarele din față ale lui Alactaga sunt mici și scurte. Iepurele de pământ le folosește când sapă, dar nu atinge pământul cu ele când sare. Săriturile se efectuează numai cu ajutorul picioarele din spateși chiar fără ajutorul unei cozi. Acesta din urmă atinge pământul doar într-o poziție calmă, când animalul se sprijină pe coadă. Scheletul piciorului are particularitatea că oasele metatarsiene ale celui de-al 2-lea, al 3-lea și al 4-lea deget de la picioare se contopesc într-un singur os. Capetele oaselor metatarsiene nu sunt topite dedesubt, iar medianul (al treilea deget) iese mai departe. Dintre falangele legate de ele, cele din mijloc sunt din nou cele mai lungi. Degetele 1 și 5 sunt vestigiale și nu ating solul. Ambele degete au aceeași lungime datorită faptului că metatarsian deget mare mai lung decât al 5-lea. Prima are 2, a doua - 3 falange.
La Dipus, specializarea a mers și mai departe: degetul 1 a dispărut fără urmă și a rămas doar un rudiment din al 5-lea. Degetul mijlociu de două ori mai slabe decât cele două exterioare și doar puțin mai lungi decât ei. Degetele sunt paralele. Degetul mijlociu este redus deoarece funcția lui este preluată de al 2-lea și al 4-lea, ale căror capete oferă sprijin în ședință și sărituri. În evoluție ulterioară, al 3-lea deget trebuie să dispară: în Dipus avem un tip de adaptare neterminat.
Centură scapulară mamifere simplificate, este format din mari omoplati, având o excrescență coracoidă și claviculă(Fig. 80, 81). În stadiul embrionar de dezvoltare și la monotremele adulte coracoid este un os independent.
Spatula situat în partea de sus a pieptului, are o formă triunghiulară și o excrescentă mare (crestă) cu acromiale un proces de atașare a mușchilor. La locația procesului coracoid și a procesului acromial al scapulei există fosa glenoidiană pentru articularea membrului anterior.
Claviculaîn formă de tijă servește la conectarea umărului și a sternului. Clavicula este cel mai dezvoltată la animalele care efectuează mișcări complexe cu membrele anterioare (lilieci, alunițe, primate, rozătoare etc.). La multe specii caracterizate prin mișcări monotone (ungulate, câini), clavicula este absentă. În general, centura membrelor anterioare este legată de scheletul axial prin mușchi și ligamente.
Brâu pelvian (Fig. 82 A) constă din pereche oase fără nume, format din articulatie iliacă, ischială și pubiană oase. Pelvis tip închis iar prin oasele iliace este ferm legat de regiune sacră coloana vertebrală. Când elementele pelvisului cresc împreună, se întâmplă fuziunea pubiană si se formeaza acetabul pentru articularea membrului posterior. În partea inferioară a osului innominat există foramenul obturator.
Scheletul membrelor pereche
Membrele mamiferelor, ca și cele ale altor vertebrate terestre, sunt de tipul cu cinci degete și reprezintă o pârghie cu trei membri, toate părțile fiind conectate mobil între ele.
Membre anterioare cuprinde umăr, antebraț și mână(Fig. 81). Osul brahial bine dezvoltat, are capul rotunjit pentru prinderea de brâul membrelor. Antebraț alcătuiește două oase - radial si ulnar, având olecranîn vârf. La joncțiunea umărului și antebrațului există articulația cotului.
Perie include trei departamente - încheietura mâinii, metacarp și falange. Încheietura este format din 8–10 elemente dispuse pe trei rânduri . Chişiţă include cinci oase (după numărul degetelor). Fiecare deget are trei falange, cu excepția primului deget, care este format din două falange. Spre deosebire de articulația intercarpiană a reptilelor și păsărilor, mobilul comun la mamifere se localizează între antebraț și mână(ca amfibienii).
Membre posterioare mamiferele au, de asemenea, trei diviziuni - coapsă, picior și picior(Fig. 82 B). Femur masiv și introdus cu capul rotunjit în acetabulul centurii pelvine. Tibia conține două oase - tibiei și fibulei. Tibia mai mare formează partea interioară a piciorului inferior, iar fibula alcătuiește partea sa exterioară.
Între coapsă și tibie există comun, acoperit rotula genunchiului, care a fost format evolutiv din tendoanele lor musculare osificate. Suprafețele oaselor articulare sunt acoperite cu formațiuni cartilaginoase. Articulația gleznei La mamifere, este situat între capetele inferioare ale tibiei și partea proximală a piciorului.

Orez. 82. Brâul pelvin (A) și membrul posterior (B) al unei vulpi:
1 – ilium, 2 – ischion, 3 – osul pubian, 4 – acetabul, 5 – femur, 6 - rotula, 7 - tibiei, 8 - peroneu, 9 - tars, 10 - metatars, 11 - falange, 12 - talus, 13 - calcaneus
Elemente tarsale picioarele sunt aranjate pe trei rânduri și formează o excrescență - calcaneului, caracteristic mamiferelor. Metatars este format din cinci oase de care sunt atașate falangele degetelor. Ca și în membrele anterioare, sunt trei în fiecare deget, cu excepția primului (interior), care are doar două falange.
Mamiferele se caracterizează printr-o aranjare specială a membrelor în raport cu corp: ele sunt situate sub corp, ceea ce contribuie la dezvoltarea unei mai mari manevrabilitati a mișcărilor. în care articulația cotuluiîndreptată posterior, iar articulația genunchiului, dimpotrivă, este orientată anterior.
Datorită diversității speciilor de mamifere, stilului lor de viață și metodelor de mișcare, lungimea relativă a membrelor, raportul elementelor constitutive, configurația oaselor, numărul degetelor etc. variază semnificativ. De exemplu, la lilieci lungimea membrului anterior, în special falangele degetelor, este mărită brusc, servind la fixarea membranei de zbor.
Dimpotrivă, membrul anterior al aluniței este scurtat, partea palmară a mâinii este lărgită, iar al șaselea deget este bine dezvoltat. La balene, mâna este alungită brusc din cauza creșterii numărului de falange ale degetelor, în urma căreia membrul a căpătat o mare asemănare cu aripioarele peștilor cu aripioare lobe. Degetele animalelor cu biomasă mare și alergare rapidă (ungulate cu degetele impare, ungulatele cu degetele pare) au fost reduse și modificate semnificativ.
Structura externă a câinelui
Câinele a fost primul animal domestic. Omul a îmblânzit-o în vremuri străvechi. Câinele a ajutat la omul primitivîn timpul vânătorii, ea i-a păzit casa. În zilele noastre sunt cunoscuți câini de lucru, de vânătoare și decorativi, precum și câinii mestiți. Câinii ajută la protejarea granițelor Patriei noastre și în timpul Marelui Războiul Patriotic a căutat răniții, a ajutat la recunoaștere și comunicații. Oamenii de știință efectuează cercetări asupra funcțiilor vitale ale corpului la câini. Câine înaintea omului spațiu vizitat.
Corpul unui câine, cum ar fi un ciobănesc sau un husky, este zvelt și musculos. Membrele sale nu sunt situate pe părțile laterale ale corpului, ca cele ale reptilelor, ci sub corp, astfel încât corpul animalului nu atinge pământul. Câinele merge, sprijinindu-se pe degete cu gheare puternice. Un gât flexibil permite o mai mare mobilitate a capului. Gura animalului este limitată de buzele mobile - superioare și inferioare. De mai sus buza superioară există un nas cu o pereche de deschideri nazale externe - nări. Ochii au pleoape bine dezvoltate. Membrana nictitante (a treia pleoapă) la câine, ca la toate mamiferele, este subdezvoltată. Dintre toate animalele, doar mamiferele au urechea externa - Pavilionul urechii. Urechea exterioară a unui câine este mare și mobilă.
Coperte de mamifere
Pielea mamiferelor este puternică și elastică; la majoritatea animalelor, bazele părului sunt situate în ea, care formează o trăsătură caracteristică acestei clase de vertebrate. linia părului . Există groși și par lung — mustaţăși altele mai scurte, mai moi - sub blană, sau subpar. Capul aspru și puternic protejează subpelul și pielea de deteriorare. Subpelul, care reține mult aer, reține bine căldura corpului. Pe lângă coloana vertebrală și sub blană, animalele dezvoltă păr mare - organe de atingere.
Părul mamiferelor, ca penele păsărilor și solzii reptilelor, este format dintr-o substanță cornoasă. De două ori pe an, un câine, la fel ca multe alte mamifere, naște - o parte din părul său cade și este înlocuit cu altele noi. Baza părului este situată în interiorul unui folicul de păr special, în care se deschid canalele firelor de păr din apropiere. glande sebacee. Secrețiile lor lubrifiază pielea și părul, făcându-le elastice și nu umezite de apă.
Figura: Structura pielii și părului unui mamifer
În pielea majorității animalelor sunt localizate și glandele sudoripare . Transpirația se evaporă de pe suprafața corpului, răcindu-l. Odată cu transpirația, excesul de sare și uree sunt, de asemenea, îndepărtate din organism; astfel, glandele sudoripare joacă rolul unor organe excretoare suplimentare.
Un câine are puține glande sudoripare și răcirea corpului se realizează prin creșterea respirației.
La capetele degetelor de mamifere se află gheare, unghii sau copite cornoase. Uneori, pe cap se dezvoltă și formațiuni cornoase (coarne la rinocer, antilope, bovine etc.) sau pe coadă (de exemplu, solzi cornos la șobolani).
Scheletul de mamifer
Scheletul mamiferelor este format din aceleași părți ca și cele ale altor vertebrate. Craniul animalelor se distinge printr-un craniu mai mare, care este asociat cu dimensiuni mari creier. Este foarte tipic ca mamiferele să aibă 7 vertebre cervicale. Atât girafele cu gâtul lung, cât și balenele au același număr de vertebre cervicale. Vertebra toracica(de obicei sunt 12-15 dintre ele) împreună cu coastele și sternul formează un puternic cufăr. Vertebrele masive ale regiunii lombare sunt articulate mobil unele cu altele. În această secțiune, trunchiul se poate îndoi și desfășura. Numărul de vertebre lombare variază între tipuri diferite(2-9), câinele are 6 dintre ele. Secțiune sacră coloana vertebrală (3-4 vertebre) fuzionează cu oasele pelvine. Numărul de vertebre din regiunea caudală (de la trei la câteva zeci) depinde de lungimea cozii.

Figura: Schelet de mamifer folosind exemplul unui câine
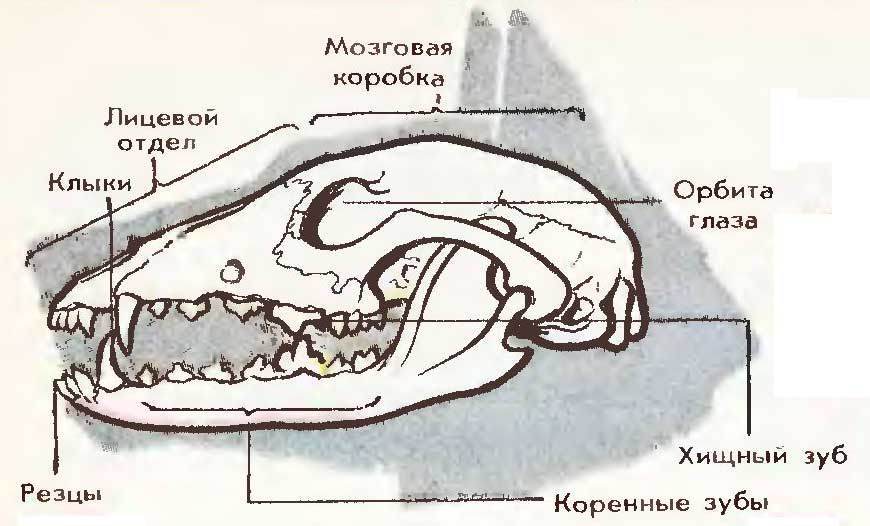
Model: Craniu de câine
Centura membrelor anterioare mamiferele este alcătuită din doi omoplați cu oase de corb atașate la ei și două clavicule. Claviculele câinelui nu sunt dezvoltate. Centura membrelor posterioare- pelvis - format din trei perechi oasele pelvine. Scheletele membrelor diferitelor specii de mamifere și reptile sunt similare, dar detaliile structurii lor sunt diferite în diferite specii și depind de condițiile de viață ale animalului.
Mușchii mamiferelor
Majoritatea mamiferelor, inclusiv câinii, au mușchi dezvoltați în special în spate, membre și brâuri. Câinele poate alerga cu salturi mari, îndoindu-și și îndreptându-și corpul, împingând alternativ de pe pământ cu picioarele din față și din spate. Mușchi puternici pus in miscare maxilarul inferior. Strângând fălcile înarmate cu dinți, câinele își ține ferm prada.

Figura: Mușchii câinelui care mișcă maxilarul inferior
Coloana vertebrală este formată din secțiunile cervicale, toracice, lombare, sacrale și caudale. A lui trăsătură caracteristică- forma platiceliala (cu suprafete plane) a vertebrelor, intre care se afla cartilaginoase discuri intervertebrale. Arcurile superioare sunt bine definite. Există șapte vertebre în regiunea cervicală, a căror lungime determină lungimea gâtului; doar lamantinul și leneșul - Choloepus hof-fmani au 6 dintre ei, iar leneșul - Bradypus 8-10. Vertebra cervicala foarte lungi la girafe si foarte scurti la cetacee, care nu au interceptare cervicala. La vertebre toracic coastele sunt articulate pentru a forma pieptul. Sternul care îl închide este plat și numai la lilieci și la speciile vizuitoare cu membrele anterioare puternice (de exemplu, alunițe) are o creastă mică (chilă), care servește ca punct de atașare muschii pectorali. În regiunea toracică sunt 9-24 (de obicei 12-15) vertebre, ultimele 2-5 vertebre toracice poartă coaste false care nu ajung la stern.În regiunea lombară sunt de la 2 până la 9 vertebre; Coastele rudimentare se contopesc cu procesele lor transversale mari. Secțiunea sacră este formată din 4-10 vertebre topite, dintre care doar primele două sunt cu adevărat sacrale, iar restul sunt caudale. Numărul de vertebre caudale libere variază de la 3 (la gibon) la 49 la șopârla cu coadă lungă.
Gradul de mobilitate al vertebrelor individuale variază. La animalele mici care alergă și cățără, este mare, de-a lungul întregii lungimi a coloanei vertebrale, astfel încât corpul lor se poate îndoi în direcții diferite și chiar se poate încolăci într-o minge. Vertebrele toracice și regiunile lombare la animalele mari, cu mișcare rapidă. La mamiferele care se deplasează pe picioarele posterioare (canguri, jerboi, săritori), cele mai mari vertebre sunt situate la baza cozii și a sacrului, iar mai departe dimensiunea lor scade succesiv. La ungulate, dimpotrivă, vertebrele și în special procesele spinoase ale acestora sunt mai mari în partea anterioară a regiunii toracice, unde de ele sunt atașați mușchii puternici ai gâtului și parțial membrele anterioare (
Articole similare
-
Cum este aranjată o biserică ortodoxă în interior?
Unde s-au rugat primii creștini? Ce sunt octogonul, transeptul și naosul? Cum este structurat un templu cu cort și de ce a fost această formă atât de populară în Rus'? Unde este cel mai înalt loc din templu și despre ce vă vor spune frescele? Ce obiecte sunt în altar? Să împărțim...
-
Venerabilul Gherasim din Vologda
Principala sursă de informații biografice despre călugărul Gherasim este „Povestea minunilor lui Gherasim din Vologda”, scrisă de un anume Toma în jurul anului 1666, cu binecuvântarea arhiepiscopului Markel de Vologda și Great Perm. Conform poveștii...
-
Sfânta Egale cu Apostolii Nina, iluminatoarea Georgiei Moaștele Sfintei Nina
În toamna anului 2016, surorile Mănăstirii Sfintei Treimi Stefano-Makhrischi Stavropegic au făcut un pelerinaj în locurile sfinte ale Georgiei. În ajunul sărbătoririi pomenirii sfântului luminator al Iveriei, vă oferim un reportaj foto despre...
-
Persoanele născute pe 8 aprilie soarta
Persoanele născute în această zi sunt extrem de active. Priviți viața ca pe o serie de provocări și intenționați să le rezolvați pe toate. Realizând abilitățile tale creative sau acționând ca șef al unei mari corporații,...
-
Ora de curs „să ne înclinăm în fața acelor ani grozavi” Scenariul pentru ora de clasă pentru 9 mai
Întocmită de un profesor de școală primară la Școala Gimnazială Nr. 1 MKOU. Ora cursului Izberbash. Scop: Crearea condițiilor necesare care să conducă la educarea sentimentelor patriotice în rândul școlarilor mai mici, formarea propriului lor civil și patriotic...
-
Formarea abilităților cognitive în școala primară
Discurs de Gusarova S.A. la ședința profesorilor pe tema: Formarea abilităților de învățare cognitivă în lecțiile din școala primară „Copilul nu vrea să ia cunoștințe gata făcute și îl va evita pe cel care îi bate cu forța în cap. Dar el de bunăvoie...