Нервный импульс. Нервные импульсы
Нейроны сообщаются между собой при помощи «нервных сообщений». Эти «сообщения» похожи на электрический ток, который бежит по проводам. Иногда, при передаче от одного нейрона к другому, эти импульсы превращаются в химические сообщения.
Нервные импульсы
Информация передается между нейронами подобно электрическому току в проводах. Эти сообщения закодированы: они представляют собой последовательность совершенно одинаковых импульсов. Сам код кроется в их частоте, то есть в числе импульсов в секунду. Импульсы передаются от клетки к клетке, от дендрита, в котором они возникают, к аксону, через который они проходят. Но есть и отличие от электрических сетей - импульсы передаются не при помощи электронов*, а при помощи более сложных частиц - ионов.
Медикаменты, влияющие на скорость импульсов
Существует множество химических препаратов, способных изменить характеристики передачи нервных импульсов. Как правило, они действуют на синаптическом уровне. Анестетики и транквилизаторы замедляют, а иногда и вообще подавляют передачу импульсов. А антидепрессанты и стимуляторы, такие как кофеин, наоборот способствуют лучшей их передаче.
С огромной скоростью
Нервные импульсы должны быстро проходить по телу. Ускорить их прохождение нейронам помогают окружающие их глиальные клетки. Они образуют оболочку нервного волокна, называемую миелиновой. В результате импульсы идут с умопомрачительной скоростью - более 400 км/час.
Химические связи
Передаваемые от нейрона к нейрону сообщения должны превращаться из электрической в химическую форму. Это связано с тем, что, несмотря на свою многочисленность, нейроны никогда не соприкасаются между собой. Но электрические импульсы не могут передаваться, если нет физического контакта. Поэтому нейроны используют для связи между собой специальную систему, называемую синапсами. В этих местах нейроны разделены узким пространством синаптической щелью. Когда электрический импульс приходит к первому нейрону, он высвобождает из синапса химические молекулы, так называемые нейромедиаторы. Эти вещества, вырабатываемые нейронами, перемещаются через синаптическую щель и попадают на специально предназначенные для них рецепторы другого нейрона. В результате возникает еще один электрический импульс.
Импульс между нейронами проходит меньше, чем за тысячную секунды.
Различие нейро-медиаторов
Мозгом вырабатывается около полусотни нейромедиаторов, которые можно подразделить на две группы. Первая состоит из тех, что инициируют возникновение нервного импульса, - их называют возбуждающими. Другие, напротив, замедляют его возникновение - это тормозящие нейромедиаторы. Стоит отметить, что в большинстве случаев нейрон выделяет только один тип нейромедиаторов. И в зависимости оттого, является ли он возбуждающим или тормозящим, нейрон по-разному воздействует на соседние нервные клетки.
Искусственная стимуляция
Отдельный нейрон или группу нейронов возможно стимулировать искусственно при помощи введенных в них электродов, направляющих электрические импульсы в точно обозначенные зоны мозга. Этот метод иногда используют в медицине, в частности для лечения больных страдающих болезнью Паркинсона Эта проявляющаяся в пожилом возрасте болезнь сопровождается дрожанием конечностей. Это дрожание может быть остановлено путем постоянной стимуляции конкретной зоны мозга.
Нейрон — микрокомпьютер
Каждый из нейронов способен принимать сотни сообщений в секунду. И, чтобы не оказаться перегруженным информацией, он должен уметь судить о степени ее значимости и делать ее предварительный анализ. Эта вычислительная деятельность происходит внутри клетки. Там складываются возбуждающие и вычитаются тормозящие импульсы. И, для того чтобы нейрон сгенерировал собственный импульс, необходимо, чтобы сумма предыдущих оказалась больше определенного значения. Если сложение возбуждающих и тормозящих импульсов не превысит этот предел, нейрон будет «молчать».
Информационные дороги
Во всем этом хитросплетении нейронов существуют прекрасно обозначенные пути. Схожие идеи, схожие воспоминания проходят, приводя всегда в действие одни и те же нейроны и синапсы. До сих пор неизвестно, как возникают и поддерживаются эти, подобные контурам электронных схем связи, но очевидно, что они существуют и что, чем они прочнее,тем они эффективнее. Часто используемые синапсы работают быстрее. Этим и объясняется то, почему мы быстрее вспоминаем вещи, которые мы видели или повторяли несколько раз. Однако эти связи возникают не навсегда. Некоторые из них могут исчезнуть, если их недостаточно использовали, а на их месте возникнуть новые. При необходимости нейроны всегда способны создавать новые связи.
Маленькие зеленые точки на фото - гормоны внутри кровеносных сосудов
Химический допинг
Когда говорят, что спортсмен использовал гормональный допинг, это значит, что он принимал гормоны либо в виде таблеток, либо вводя их непосредственно в кровь. Гормоны бывают естественными или искусственными. Самые распространенные - гормоны роста и стероиды, за счет которых мышцы становятся больше и сильнее, а также эритропоэтин - гормон, ускоряющий доставку питательных веществ к мышцам.
Мозг способен производить миллионы операций за доли секунды.
На мозг работают гормоны
Для обмена информацией мозгом используется и другой инструмент - гормоны . Эти химические соединения частично производятся самим мозгом в группе нейронов, расположенных в гипоталамусе. Эти гормоны контролируют производство иных, вырабатываемых в других частях тела в эндокринных железах. Они действуют иначе, чем нейромедиаторы, которые фиксируются непосредственно на нейронах и переносятся с кровью к отдаленным от мозга органам тела, таким как груди, яичники, мужские семенники, почки. Закрепляясь на их рецепторах, гормоны вызывают различные физиологические реакции. Они, например, способствуют росту костей и мышц, управляют чувством голода и жажды и, конечно, влияют на сексуальную активность.
НЕРВНЫЙ ИМПУЛЬС
- волна возбуждения, к-рая
распространяется по нервному волокну и служит для передачи информации от периферич.
рецепторных (чувствительных) окончаний к нервным центрам, внутри центр. нервной
системы и от неё к исполнительным аппаратам - мышцам и железам. Прохождение
Н. и. сопровождается переходными электрич. процессами, к-рые можно зарегистрировать
как внеклеточными, так и внутриклеточными электродами.
Генерацию, передачу и переработку Н. и. осуществляет
нервная система. Осн. структурным элементом нервной системы высших организмов
является нервная клетка, или нейрон, состоящий из тела клетки и многочисл. отростков
- дендритов (рис. 1). Один из отростков у нериферич. нейронов имеет большую
длину - это нервное волокно, или аксон, протяжённость к-рого ~ 1 м, а толщина
от 0,5 до 30 мкм. Различают два класса нервных волокон: мякотные (мие-линизированные)
и безмякотные. У мякотных волокон имеется миелиновая оболочка, образованная
спец. мембраной, к-рая подобно изоляции накручивается на аксон. Протяжённость
участков сплошной миелиновой оболочки составляет от 200 мкм до 1 мм, они прерываются
т. н. перехватами Ранвье шириной 1мкм. Миелиновая оболочка играет роль изоляции;
нервное волокно на этих участках пассивно, электрически активна только мембрана
в перехватах Ранвье. Безмякотные волокна не имеют изолир. участков; их структура
однородна по всей длине, а мембрана обладает электрич. активностью по всей поверхности.
Нервные волокна заканчиваются на телах или ден-дритах
др. нервных клеток, но отделены от них проме-
жутком шириной ~ 10 нм. Эта область контакта
двух клеток наз. синапсом. Входящая в синапс мембрана аксона наз. пресинаптической,
а соответствующая мембрана дендритов или мышцы - пост-синаптической (см. Клеточные
структуры)
.
В нормальных условиях по нервному волокну постоянно
бегут серии Н. и., возникающих на дендритах или теле клетки и распространяющихся
по аксону в направлении от тела клетки (аксон может проводить Н. и. в обоих
направлениях). Частота этих периодич. разрядов несёт информацию о силе вызвавшего
их раздражения; напр., при умеренной активности частота ~ 50-100 импульсов/с.
Существуют клетки, к-рые разряжаются с частотой ~ 1500 импульсов/с.
Скорость распространения Н. и. u
зависит от типа нервного волокна и его диаметра d
, u
~ d
1/2 . В тонких волокнах нервной системы человека u
~ 1 м/с, а в толстых волокнах u
~ 100-120 м/с.
Каждый Н. и. возникает в результате раздражения
тела нервной клетки или нервного волокна. Н. и. всегда имеет одни и те же характеристики
(форму и скорость) независимо от силы раздражения, т. е. при подпороговом раздражении
Н. и. не возникает совсем, а при надпороговым - имеет полную амплитуду.
После возбуждения наступает рефракторный период, в течение к-рого возбудимость нервного волокна снижена. Различают абс. рефракторный период, когда волокно нельзя возбудить никакими раздражителями, и относит. рефракторный период, когда возбуждение возможно, но его порог оказывается выше нормы. Абс. рефракторный период ограничивает сверху частоту передачи Н. и. Нервное волокно обладает свойством аккомодации, т. е. привыкает к постоянно действующему раздражению, что выражается в постепенном повышении порога возбудимости. Это приводит к снижению частоты Н. и. и даже к их полному исчезновению. Если сила раздражения нарастает медленно, то возбуждения может не произойти даже после достижения порога.
Рис.1. Схема строения нервной клетки .
Вдоль нервного волокна Н. и. распространяется
в виде волны электрич. потенциала. В синапсе происходит смена механизма распространения.
Когда Н. и. достигает пресинаптич. окончания, в синаптич. щель выделяется активное
хим. вещество - м е д и а т о р. Медиатор диффундирует через синаптич. щель
и меняет проницаемость постсинаптич. мембраны, в результате чего на ней возникает
потенциал, вновь генерирующий распространяющийся импульс. Так действует хим.
синапс. Встречается также электрич. синапс, когда след. нейрон возбуждается
электрически.
Возбуждение Н. и
. Физ. представления о
появлении электрич. потенциалов в клетках основаны на т. н. мембранной теории.
Клеточные мембраны разделяют растворы электролита разной концентрации и обладают
из-бират. проницаемостью для нек-рых ионов. Так, мембрана аксона представляет
собой тонкий слой липидов и белков толщиной ~ 7 нм. Её электрич. сопротивление
в состоянии покоя ~ 0,1 Ом. м 2 , а ёмкость ~ 10 мф/м 2 .
Внутри аксона высока концентрация ионов К + и мала концентрация ионов
Na + и Сl - , а в окружающей среде - наоборот.
В состоянии покоя мембрана аксона проницаема
для ионов К + . Из-за разницы концентраций C 0 K
во внеш. и С во внутр. растворах на мембране устанавливается калиевый мембранный
потенциал
![]()
где Т
- абс. темп-pa, е
-
электрона. На мембране аксона действительно наблюдается потенциал покоя ~ -60
мВ, соответствующий указанной ф-ле.
Ионы Na + и Сl - проникают
через мембрану. Для поддержания необходимого неравновесного распределения ионов
клетка использует систему активного транспорта, на работу к-рой расходуется
клеточная энергия. Поэтому состояние покоя нервного волокна не является термодинамически
равновесным. Оно стационарно благодаря действию ионных насосов, причём мембранный
потенциал в условиях разомкнутой цепи определяется из равенства нулю полного
электрич. тока.
Процесс нервного возбуждения развивается следующим
образом (см. также Биофизика
).Если пропустить через аксон слабый импульс
тока, приводящий к деполяризации мембраны,
то после снятия внеш. воздействия потенциал монотонно возвращается к исходному
уровню. В этих условиях аксон ведёт себя как пассивная электрич. цепь, состоящая
из конденсатора и пост. сопротивления.
Рис. 2. Развитие потенциала действия в нервном
во
локне: а
- подпороговое
(1
) и надпороговое (2
) раздражения;
б
-мембранный отклик; при над-пороговом раздражении проявляется полный
потен
циал действия; в
- ионный
ток, протекающий через
мембрану при
возбуждении; г
- аппроксимация
ионного
тока в простой аналитической модели
.
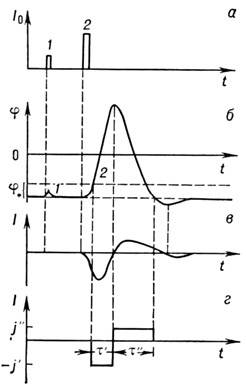
Если импульс тока превышает нек-рую пороговую
величину, потенциал продолжает изменяться и после выключения возмущения; потенциал
становится положительным и только потом возвращается к уровню покоя, причём
вначале даже несколько проскакивает его (область гиперполяризации, рис. 2).
Отклик мембраны при этом не зависит от возмущения; этот импульс наз. потенциалом
действия. Одновременно через мембрану течёт ионный ток, направленный сначала
внутрь, а потом наружу (рис. 2, в
).
Феноменологич. истолкование механизма возникновения
Н. и. было дано А. Л. Ходжкином (A. L. Hodg-kin) и А. Ф. Хаксли (A. F. Huxley)
в 1952. Полный ионный ток слагается из трёх составляющих: калиевого, натриевого
и тока утечки. Когда потенциал мембраны сдвигается на пороговую величину j*
(~ 20мВ), мембрана становится проницаемой для ионов Na + . Ионы Na +
устремляются внутрь волокна, сдвигая мембранный потенциал, пока он не достигнет
величины равновесного натриевого потенциала:
![]()
составляющего ~ 60 мВ. Поэтому полная амплитуда
потенциала действия достигает ~ 120 мВ. К моменту достижения макс. потенциала
в мембране начинает развиваться калиевая (и одновременно уменьшаться натриевая)
проводимость. В результате натриевый ток сменяется
на калиевый, направленный наружу. Этот ток соответствует уменьшению потенциала
действия.
Установлены эмпирич. ур-ния для описания натриевого
и калиевого токов. Поведение мембранного потенциала при пространственно однородном
возбуждении волокна определяется ур-нием:
где С
- ёмкость мембраны, I
- ионный
ток, слагающийся из калиевого, натриевого и тока утечки. Эти токи определяются
пост. эдс j K ,
j Na
и j l
и проводимостями g
K , g
Na и g l:
Величину g l
считают постоянной,
проводимости g
Na и g
K описывают с
помощью параметров m
, h
и п:
g
Na , g
K -
постоянные; параметры т, h
и п
удовлетворяют линейным ур-ниям
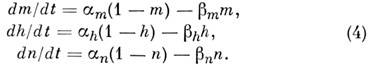
Зависимость коэф. a
и b от мембранного
потенциала j
(рис. 3) выбирают из условия наилучшего совпадения
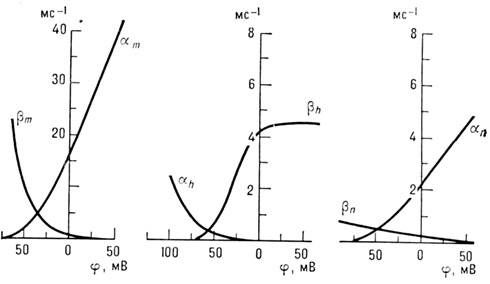
Рис. 3. Зависимость коэффициентов
a
и
b
от мембран
ного потенциала
.
расчётных и измеряемых кривых I
(t
).
Этими же соображениями вызван выбор параметров. Зависимость стационарных значений
т, h
и п
от мембранного потенциала приведена на рис. 4. Существуют
модели с большим числом параметров. Т. о., мембрана нервного волокна представляет
собой нелинейный ионный проводник, свойства к-рого существенно зависят от электрич.
поля. Механизм генерации возбуждения изучен плохо. Ур-ния Ходжкина -Хаксли дают
лишь удачное эмпирич. описание явления, за к-рым нет конкретной физ. модели.
Поэтому важной задачей является изучение механизмов протекания электрич. тока
через мембраны, в частности через управляемые элект- рич. полем ионные каналы.
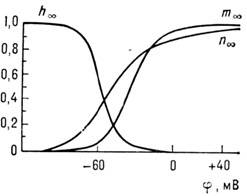
Рис. 4. Зависимость стационарных значений
т, h
и п
от мембранного
потенциала
.
Распространение Н. и
. Н. и. может распространяться
вдоль волокна без затухания и с пост. скоростью. Это связано с тем, что необходимая
для передачи сигнала энергия не поступает из единого центра, а черпается на
месте, в каждой точке волокна. В соответствии с двумя типами волокон существуют
два способа передачи Н. и.: непрерывный и сальтаторный (скачкообразный), когда
импульс движется от одного перехвата Ранвье к другому,
перепрыгивая через области миелиновой изоляции.
В случае немиелинизир. волокна распределение
мембранного потенциала j(x,
t
)определяется ур-нием:
где С
- ёмкость мембраны, приходящаяся
на единицу длины волокна, R
- сумма продольных (внутриклеточного и внеклеточного)
сопротивлений на единицу длины волокна, I
- ионный ток, протекающий через
мембрану волокна единичной длины. Электрич. ток I
является функционалом
от потенциала j,
к-рый зависит от времени t
и координаты х
. Эта зависимость определяется
ур-ниями (2) -(4).
Вид функционала I
специфичен для биологически
возбудимой среды. Однако ур-ние (5), если отвлечься от вида I
, имеет
более общий характер и описывает многие физ. явления, напр. процесс горения.
Поэтому передачу Н. и. уподобляют горению порохового шнура. Если в бегущем пламени
процесс поджигания осуществляется за счёт , то в Н. и. возбуждение
происходит при помощи т. н. локальных токов (рис. 5).
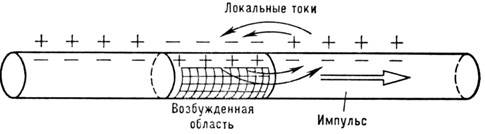
Рис. 5. Локальные токи, обеспечивающие распростра
нение
нервного импульса
.
Ур-ния Ходжкина - Хаксли для распространения
Н. и. решались численно. Полученные решения вместе с накопленными эксперим.
данными показали, что распространение Н. и. не зависит от деталей процесса возбуждения.
Качеств. картину распространения Н. и. можно получить при помощи простых моделей,
отражающих лишь общие свойства возбуждения. Такой подход позволил рассчитывать
скорость и форму Н. и. в однородном волокне, их изменение при наличии неоднород-ностей
и даже сложные режимы распространения возбуждения в активных средах, напр. в
сердечной мышце. Существует неск. матем. моделей подобного рода. Простейшая
из них такова. Ионный ток, протекающий через мембрану при прохождении Н. и.,
является знакопеременным: вначале он течёт внутрь волокна, а потом наружу. Поэтому
его можно аппроксимировать кусочно-постоянной ф-цией (рис. 2, г
). Возбуждение
происходит, когда мембранный потенциал сдвигается на пороговую величину j*.
В этот момент возникает ток, направленный внутрь волокна и равный по модулю
j"
. Спустя время t"
ток меняется на противоположный, равный j
"". Эта фаза продолжается
в течение времени ~ t"".
решение ур-ния (5) можно найти как ф-цию переменной t = х/
u, где u
- скорость распространения Н. и. (рис. 2, б)
.
В реальных волокнах время t"
достаточно велико, поэтому только оно определяет скорость u, для к-рой справедлива ф-ла: ![]() . Учитывая, что j
" ~ ~d, R ~ d
2 и С
~
d
, где d
- диаметр волокна, находим в согласии с экспериментом,
что u ~ d
1/2 . С помощью кусочно-постоянной аппроксимации находят форму потенциала
действия.
. Учитывая, что j
" ~ ~d, R ~ d
2 и С
~
d
, где d
- диаметр волокна, находим в согласии с экспериментом,
что u ~ d
1/2 . С помощью кусочно-постоянной аппроксимации находят форму потенциала
действия.
Ур-ние (5) для распространяющегося Н. и. в действительности
допускает два решения. Второе решение оказывается неустойчивым; оно даёт Н.
и. со значительно меньшей скоростью и амплитудой потенциала. Наличие второго,
неустойчивого, решения имеет аналогию в теории горения. При распространении
пламени с боковым теплоотводом также возможно возникновение неустойчивого режима.
Простую аналитич. модель Н. и. можно усовершенствовать, учитывая дополнит. детали.
При изменении сечения и при ветвлении нервных
волокон прохождение Н. и. может быть затруднено или даже полностью блокировано.
В расширяющемся волокне (рис. 6) скорость импульса по мере приближения к расширению
убывает, а после расширения начинает расти, пока не выйдет на новое стационарное
значение. Замедление Н. и. тем сильнее, чем больше разница в сечениях. При достаточно
большом расширении Н. и. останавливается. Существует критич. расширение волокна,
к-рое задерживает Н. и.
При обратном движении Н. и. (из широкого волокна
в узкое) блокирования не происходит, но изменение скорости носит противоположный
характер. При подходе к сужению скорость Н. и. увеличивается, а затем начинает
спадать до нового стационарного значения. На графике скорости (рис., 6а
)
получается своего рода петля .
Рие. 6. Прохождение нервных импульсов по расширя
ющемуся
волокну: а
- изменение скорости импульса в
зависимости
от его направления; б
-схематическое
изображение
расширяющегося волокна
.
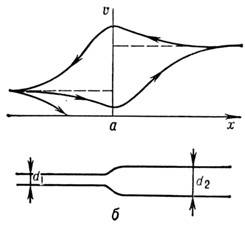
Другой тип неоднородности - ветвление волокон.
В узле ветвления возможны разл. варианты прохождения и блокирования импульсов.
При несинхронном подходе Н. и. условие блокирования
зависит от временного сдвига. Если временной сдвиг между импульсами мал, то
они помогают друг другу проникнуть в широкое третье волокно. Если сдвиг достаточно
велик, то Н. и. мешают друг другу. Связано это с тем, что Н. и., подошедший
первым, но не сумевший возбудить третье волокно, частично переводит узел в рефракторное
состояние. Кроме того, возникает эффект синхронизации: по мере приближения Н.
и. к узлу их запаздывание друг относительно друга уменьшается.
Взаимодействие Н. и
. Нервные волокна в
организме объединены в пучки или нервные стволы, образующие подобие многожильного
кабеля. Все волокна в пучке представляют собой самостоят. линии связи, но имеют
один общий "провод" - межклеточную жидкость. Когда по любому из
волокон бежит Н. и., он создаёт в межклеточной жидкости электрич. поле, к-рое
влияет на мембранный потенциал соседних волокон. Обычно такое влияние пренебрежимо
мало и линии связи работают без взаимных помех, но оно проявляется в пато-логич.
и искусств. условиях. Обрабатывая нервные стволы спец. хим. веществами, удаётся
наблюдать не только взаимные помехи, но и передачу возбуждения в соседние волокна.
Известны эксперименты по взаимодействию двух
нервных волокон, помещённых в ограниченный объём внеш. раствора. Если по одному
из волокон бежит Н. и., то одновременно изменяется возбудимость второго волокна.
Изменение проходит три стадии. Вначале возбудимость второго волокна падает (повышается
порог возбуждения). Это уменьшение возбудимости опережает потенциал действия,
бегущий по первому волокну, и длится примерно до тех пор, пока потенциал в первом
волокне не достигнет максимума. Затем возбудимость растёт, эта стадия совпадает
по времени с процессом уменьшения потенциала в первом волокне. Возбудимость
ещё раз уменьшается, когда в первом волокне происходит небольшая гиперполяризация
мембраны.
При одноврем. прохождении Н. и. по двум волокнам
иногда удавалось достигнуть их синхронизации. Несмотря на то что собств. скорости
Н. и. в разных волокнах различны, при их
одноврем. возбуждении мог возникнуть коллективный Н. и. Если собств. скорости
были одинаковы, то коллективный импульс имел меньшую скорость. При заметном
отличии собств. скоростей коллективная скорость имела промежуточное значение.
Синхронизоваться могли лишь Н. и., скорости к-рых отличались не слишком сильно.
Матем. описание этого явления даётся системой
ур-ний для мембранных потенциалов двух параллельных волокон j 1
и j 2:
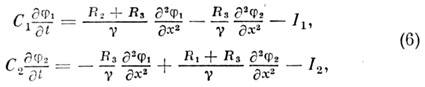
где R
1 и R
2
- продольные сопротивления первого и второго волокон, R
3
- продольное сопротивление внешней среды, g
= R
1 R
2 + R
1 R
3
+ R
2 R
3 . Ионные токи I
1
и I
2 можно описать той или иной моделью нервного возбуждения.
При использовании простой аналитич. модели решение
приводит к след. картине. Когда возбуждается одно волокно, в соседнем наводится
знакопеременный мембранный потенциал: вначале волокно гиперполяри-зуется, затем
деполяризуется и, наконец, ещё раз ги-перполяризуется. Эти три фазы соответствуют
понижению, повышению и новому понижению возбудимости во-локна. При нормальных
значениях параметров сдвиг мембранного потенциала во второй фазе в сторону деполяризации
не достигает порога, поэтому передачи возбуждения в соседнее волокно не происходит.
При одноврем. возбуждении двух волокон система (6) допускает совместное автомодельное
решение, к-рое соответствует двум Н. и., движущимся с одинаковой скоростью на
пост. расстоянии друг от друга. Если впереди находится медленный Н. и., то он
притормаживает быстрый импульс, не выпуская его вперёд; оба движутся с относительно
малой скоростью. Если же впереди находится быстрый II. и., то он подтягивает
за собой медленный импульс. Коллективная скорость оказывается близкой к собств.
скорости быстрого импульса. В сложных нейронных структурах возможно появление
автоволи
.
Возбудимые среды
. Нервные клетки в организме
объединены в нейронные сети, к-рые в зависимости от частоты ветвления волокон
разделяют на редкие и густые. В редкой сети отд. элементы возбуждаются независимо
друг от друга и взаимодействуют только в узлах ветвления, как описано выше.
В густой сети возбуждение охватывает сразу много
элементов, так что их детальная структура и способ соединения между собой оказываются
несущественными. Сеть ведёт себя как непрерывная возбудимая среда, параметры
к-рой определяют возникновение и распространение возбуждения.
Возбудимая среда может быть трёхмерной, хотя
чаще её рассматривают как двумерную поверхность. Возбуждение, возникшее в к--л.
точке поверхности, распространяется во все стороны в виде кольцевой волны. Волна
возбуждения может огибать препятствия, но не может от них отражаться, не отражается
она и от границы среды. При столкновении волн между собой происходит их взаимное
уничтожение; пройти друг сквозь друга эти волны не могут из-за наличия позади
фронта возбуждения рефракторной области.
Примером возбудимой среды является сердечный
нервно-мышечный синцитий - объединение нервных и мышечных волокон в единую проводящую
систему, способную передавать возбуждение в любом направлении. Нервно-мышечные
синцитии сокращаются синхронно, подчиняясь волне возбуждения, к-рую посылает
единый управляющий центр - водитель ритма. Единый ритм иногда нарушается, возникают
аритмии. Один из таких режимов наз. трепетанием предсердий: это автономные сокращения,
вызванные циркуляцией возбуждения вокруг препятствия, напр.
верхней или нижней вены. Для возникновения подобного режима периметр препятствия
должен превышать длину волны возбуждения, равную в предсердии человека ~ 5 см.
При трепетании происходит пе-риодич. сокращение предсердий с частотой 3-5 Гц.
Более сложный режим возбуждения представляет собой фибрилляция желудочков сердца,
когда отд. элементы сердечной мышцы начинают сокращаться без внеш. команды и
без связи с соседними элементами с частотой ~ 10 Гц. Фибрилляция приводит к
прекращению циркуляции крови.
Возникновение и поддержание спонтанной активности
возбудимой среды неразрывно связаны с возникновением источников волн. Простейший
источник волн (группа спонтанно возбуждающихся клеток) может обеспечить периодич.
пульсацию активности, так устроен водитель ритма сердца.
Источники возбуждения могут возникать и за счёт
сложной пространств. организации режима возбуждения, напр. ревербератор типа
вращающейся спиральной волны, появляющийся в простейшей возбудимой среде. Другой
вид ревербератора возникает в среде, состоящей из элементов двух типов с разными
порогами возбуждения; ревербератор периодически возбуждает то одни, то другие
элементы, меняя при этом направление своего движения и порождая .
Третий вид источника - ведущий центр (источник
эха), к-рый появляется в среде, неоднородной по реф-ракторности или порогу возбуждения.
В этом случае на неоднородности возникает отражённая волна (эхо). Наличие подобных
источников волн приводит к появлению сложных режимов возбуждения, исследуемых
в теории автоволн.
Лит.: Ходжкин А., Нервный импульс , пер. с англ., М., 1965; Катц Б., Нерв, мышца и синапс, пер. с англ., М., 1968; Ходоров Б. И., Проблема возбудимости, Л., 1969; Тасаки И., Нервное возбуждение, пер. с англ., М., 1971; Маркин В. С., Пастушенко В. Ф., Чизмад-жев Ю. А., Теория возбудимых сред, М., 1981. В. С. Маркин .
Изучение природы нервного импульса было связано с особыми трудностями, так как при прохождении импульса по нерву никаких видимых изменений не происходит. Лишь недавно, с развитием микрохимических методов, удалось показать, что во время проведения импульса нерв расходует больше энергии, потребляет больше кислорода и выделяет больше углекислоты, чем в состоянии покоя. Это указывает на то, что в проведении импульса, в восстановлении исходного состояния после проведения или в обоих этих процессах участвуют окислительные реакции.
Когда примерно 100 лет назад было установлено, что нервный импульс сопровождается определенными электрическими явлениями, возникло мнение, что сам импульс представляет собой электрический ток. В то время было известно, что электрический ток распространяется очень быстро, и поэтому высказывалось мнение, что скорость распространения нервного импульса слишком велика, чтобы ее можно было измерить. Десять лет спустя Гельмгольц измерил скорость проведения импульса, раздражая нерв, идущий к мышце, на различных расстояниях от мышцы и измеряя время, протекавшее между раздражением и сокращением. Таким способом он показал, что нервный импульс распространяется гораздо медленнее электрического - в нервах лягушки со скоростью около 30 м/сек. Это, конечно, свидетельствовало о том, что нервный импульс не есть электрический ток, подобный току в медном проводе. Кроме того, мертвый или раздавленный нерв все еще проводит ток, но не проводит нервных импульсов, и, раздражаем ли мы нерв током, прикосновением, приложением тепла или химическими факторами, возникающий при этом импульс распространяв "я со скоростью одного и того же порядка. Из этого мы заключаем, что нервный импульс представляет собой не электрический ток, а электрохимическое возмущение в нервном волокне. Вызванное раздражителем возмущение в одном участке нервного волокна вызывает такое же возмущение в соседнем участке и так далее до тех пор, пока импульс не дойдет до конца волокна. Таким образом, передача импульса подобна горению бикфордова шнура: от теплоты, выделяющейся при горении одного участка шнура, загорается следующий участок и т. д. В нерве роль теплоты выполняют электрические явления, которые, возникнув в одном участке, стимулируют следующий.
Передача нервного импульса сходна с горением бикфордова шнура и в некоторых других отношениях. Скорость горения шнура не зависит от количества тепла, затраченного при его зажигании, если только этого тепла достаточно, чтобы шнур загорелся. Не имеет значения и метод зажигания. Так же обстоит дело и с нервом. Нерв не будет реагировать, пока к нему не будет приложено раздражение определенной минимальной силы, но дальнейшее увеличение силы раздражения не заставит импульс распространяться быстрее. Это обусловлено тем, что энергию для проведения импульса доставляет сам нерв, а не раздражитель. Описанное явление отражено в законе «все или ничего»: нервный импульс не зависит от природы и силы вызвавшего его раздражителя, если только раздражитель обладает достаточной силой, чтобы вызвать появление импульса. Хотя скорость проведения не зависит от силы раздражителя, она зависит от состояния нервного волокна, и различные вещества могут замедлять передачу импульса или делать ее невозможной.
Сгоревший шнур нельзя использовать вторично, нервное же волокно способно восстанавливать свое исходное состояние и передавать другие импульсы. Оно, однако, не может проводить их непрерывно: после проведения одного импульса проходит определенное время, прежде чем волокно сможет передавать второй импульс. Этот промежуток времени, называемый рефрактерным периодом, продолжается от 0,0005 до 0,002 сек. В это время происходят химические и физические изменения, в результате которых волокно возвращается в первоначальное состояние.
Насколько нам известно, импульсы, передаваемые нейронами всех типов - двигательными, чувствительными или вставочными, в основном сходны между собой. То, что один
импульс вызывает ощущение света, другой - ощущение звука, третий - мышечное сокращение, а четвертый стимулирует секреторную деятельность железы, всецело зависит от природы тех структур, к которым приходят импульсы, а не от каких-либо особенностей самих импульсов.
Хотя нервное волокно можно стимулировать в любой его точке, в нормальных условиях возбуждение вызывается только на одном его конце, от которого импульс идет вдоль волокна до его другого конца1. Соединение между последовательными нейронами называется синапсом. Нервный импульс передается с кончика аксона одного нейрона на дендрит следующего через синаптическое соединение путем выделения у кончика аксона определенного вещества. Это вещество вызывает появление нервного импульса в дендрите следующего аксона. Передача возбуждения через синапс происходит значительно медленнее, чем передача его по нерву. В нормальных условиях импульсы проходят только в одном направлении: в чувствительных нейронах они идут от органов чувств к спинному и головному мозгу, а в двигательных - от головного и спинного мозга к мышцам и железам. Направление определяется синапсом, так как только кончик аксона способен выделять вещество, стимулирующее другой нейрон. Каждое отдельное нервное волокно может проводить импульс в обоих направлениях; при электрическом раздражении волокна где-либо в середине возникают два импульса, один из которых идет в одном направлении, а другой - в другом (эти импульсы можно обнаружить соответствующими электрическими приборами),. но лишь тот из них, который идет по направлению к кончику аксона, может стимулировать следующий нейрон в цепи. Импульс, идущий к дендриту, «остановится», достигнув его конца.
Химические и электрические процессы, с которыми связана передача нервного импульса, во многом сходны с процессами, происходящими при мышечном сокращении. Но проводящий импульсы нерв расходует очень мало энергии по сравнению с сокращающейся мышцей; теплота, образующаяся при раздражении нерва в течение 1 мин, в расчете на 1 г ткани эквивалентна энергии, выделяющейся при окислении 0,000001 г гликогена. Это " означает, что если бы нерв содержал в качестве источника энергии лишь 1% гликогена, его можно было бы стимулировать непрерывно в течение недели и запас гликогена не был бы исчерпан. При достаточном снабжении кислородом нервные волокна практически неутомляемы. Какова бы ни была природа «умственного утомления», это не может быть настоящим утомлением нервных волокон. Ссылки по теме
Роль ацетилхолина
Нервно-мышечное соединение, было изучено весьма детально, и это дало много сведений, полезных для понимания процесса химической передачи в нейронных синапсах вообще. Его функция состоит в том, чтобы передавать импульсы с относительно очень мелких двигательных нервных окончаний на крупное мышечное волокно и заставлять его сокращаться. В большинстве мионевральных соединений в мышцах позвоночных за каждым нервным импульсом следует такой же импульс в мышечных волокнах, который быстро распространяется в обе стороны по направ-лению к сухожилиям, обеспечивая достаточно синхронную активацию сократимых белков от одного конца волокна до другого. Таким образом, мионевральное соединение позвоночного служит гораздо более простой цели, чем центральные синапсы нейронов или периферические возбудительные и тормозные нервно-мышечные контакты у ракообразных, где происходит интеграция конвергирующих сигналов и где эффект одного нервного импульса лежит обычно значительно ниже порога возбуждения эффектор- ной клетки. Грубо говоря, нервно-мышечное соединение позвоночного играет роль простого реле. Можно рассматривать его также как устройство для согласования им- цедансов - род преобразователя импульсов, который дает на выходе электрический ток, достаточный для того, чтобы поднять выше порога потенциал мышечной мембраны, обладающей малым импедансом.
Как уже было отмечено, на основании одних лишь структурных данных можно сказать, что существование эффективной электрической кабельной связи между нер« вом и мышцей крайне маловероятно. В скелетной мышце лягушки миелиновое нервное волокно распадается на целый «куст» концевых безмякотных веточек толщиной примерно 1,5 мк, которые проходят в мелких бороздках поверхности мышечного волокна на протяжении около 100 мк. На всем этом концевом участке нерв образует си- наптическое соединение с мышечным волокном. Во многих местах внутри нервных окончаний электронный микроскоп позволяет видеть обычные скопления пузырьков диаметром 500 А (фиг. 27). Пресинаптическая и постсинапти- ческая мембраны разделены межклеточным промежутком, содержащим базальную мембрану. Поверхность мышечного волокна образует правильный ряд складок, расположенных под прямым углом к концевой веточке нерва.
Посмотрим, каковы возможности электрической передачи импульса через типичное мионевральное соединение в одном из самых толстых мышечных волокон лягушки (150 мк в поперечнике). Для короткого импульса тока, например такого, какой создается потенциалом действия нерва, сопротивление (входной импеданс) мышечного волокна составляет менее 50 000 ом. Для того чтобы возбудить волокно, его мембранный потенциал покоя нужно понизить с 90 мв примерно до 50 мв, а для этого требуется импульс тока силой несколько менее 10-6 а. (Позже мы увидим, что при химической передаче во многих концевых пластинках создается ток силой 2-3-10"6 а, и, таким образом, процесс обладает достаточной «резервной мощностью».)
Какую силу тока могут обеспечить нервные окончания при электрической передаче? Будем считать, что суммарная длина / синаптических разветвлений нерва у одной концевой пластинки равна примерно 1 мм, а «синаптиче- ская поверхность» (обращенная к мышечному волокну) вычисляется по формуле ир/ ж 2,3-10"5 см2. Если нервная мембрана создает направленный наружу импульс тока плотностью порядка 1 а!"см2 (обычная величина для безмя- котного нерва и для мышцы), то это даст суммарный ток 2,3-10~8 а. Если бы даже весь этот ток входил в мышечное волокно в синаптической области (что могло бы произойти в случае непрерывности протоплазмы между нервом и мышцей) и, таким образом, мог деполяризовать мышечную мембрану, то и ТОГДа он мог бы изменить потенциал всего лишь::ШванновскаИ"
"¦Нервное ¦Шнчаийе
}~~Тмк" "^Мышечное во/юкно»;
Мышечное волокно
Шванновская клетка
Нервное окончание
Фиг. 27. Схемы синаптических структур (масштаб показан лишь приблизительно),
I. Межнейронный синапс: А - строение пресинаптического окончания; Б - расположение синапса на теле нейрона. II. Нервно- мышечное соединение лягушки: А -отдельный участок соединения; Б - общее расположение окончаний двигательного аксона на мышечном волокне (участок, изображенный на схеме А, указан не-большим прямоугольником); В - схематический рисунок, сделанный по электронной микрофотографии продольного среза мышечного волокна: I - концевая мембрана аксона; 2 - «базальная мембрана», лежащая в промежутке между нервным и мышечным волокнами; 3- складчатая постсинаптическая мембрана мышечного волокна.
на 1-2 мв. Однако в действительности из-за наличия межклеточного промежутка шириной более 500 А и им-педанса постсинаптической мембраны (фиг. 26 и 27) условия для распространения тока значительно хуже. Если удельное сопротивление промежутка имеет величину порядка 100 ом-см, то изменение потенциала в нем составит всего лишь 40 мкв. Тогда ток, входящий в мышечное волокно, будет ослаблен до величины порядка Ю-11 а и сможет изменить его мембранный потенциал лишь на ничтожную величину (менее 1 мкв).
Несмотря на весьма ориентировочный характер приведенных расчетов, ясно, что отсутствие структурной непрерывности в рассмотренном нами синапсе делает электрическую кабельную передачу крайне маловероятной; им- педансы нервных окончаний и мышечного волокна настолько «не согласованы», что даже в случае непрерывности цитоплазмы трудно было бы представить себе систему, менее приспособленную для электрической передачи сигналов!
Было сделано несколько попыток проверить это в экс-перименте. Если на нерв вблизи его синапса воздействовать подпороговым током, он не вызывает заметного изменения местного потенциала в мышечном волокне (см. также стр. 169). Когда возникает импульс, он распространяется до окончаний немиелинизированных веточек и возбуждает импульс в мышечном волокне. Если поместить микроэлектрод точно против синаптического контакта между нервом и мышцей, то можно зарегистрировать два отдельных электрических изменения (фиг. 28): сначала электри-ческий ток, генерируемый импульсом в концевом разветвлении аксона, а затем, спустя 0,5-0,8 меек, еще один такой же ток, который возникает в постсинаптической мембране . Эти два события не только разделены во времени; постсинаптическое изменение можно, кроме того, избирательно снять либо с помощью определенных ве-ществ, например кураре, либо путем понижения концентрации кальция или повышения концентрации магния. Обнаруженная задержка в местах мионеврального контакта - явление постоянное и очень важное: оно ясно указывает на отсутствие электрической непрерывности в синапсе. Очевидно, кабельный процесс в пресинаптическом
Мышечное волокно
г-0-
Артефакт Импульс,
раздражения пришедший
I о нервное
і окончание
3
І -
11111111
Время, мсек »¦
Фиг. 28. Точечное отведение от поверхности нервно-мышечного соединения.
При недостатке кальция и повышенной концентрации магния постсинаптическое отклонение может отсутствовать («нулевая» реакция 1) или иметь вид «квантовой» реакции 2. В нормальных условиях происходит значительно большее отклонение 3, приводящее к мышечному спайку.
окончании прекращается, и какой-то иной, неэлектрический процесс служит промежуточным звеном между прибытием нервного импульса и возникновением электрического сигнала в мышечном волокне.
Химическая природа этого промежуточного процесса была раскрыта в работе сэра Генри Дэйла и его сотр. , которые показали, что в результате раздражения двигательного нерва выделяется ацетилхолин и что это вещество оказывает очень сильное стимулирующее действие на скелетную мышцу. Для того чтобы установить роль ацетилхолина как химического передатчика, или медиатора, необходимо было показать: 1) что он выделяется из какого-то пресинаптического участка, 2) что он действует в каком-то постсинаптическомучастке и 3) что количество его, выделяемое нервом, достаточно для инициации импульса в мышце.
Нервная система человека
Задания с выбором одного верного ответа.
А1. Нервная регуляция функций в теле человека осуществляется с помощью:
1) электрических импульсов,
2) механических раздражений,
3) гормонов,
4) ферментов.
А2. Структурной и функциональной единицей нервной системы считают:
1) нейрон,
2) нервную ткань,
3) нервные узлы,
4) нервы.
А3. Основу нервной деятельности человека и животных составляет:
1) мышление,
2) рассудочная деятельность,
3) возбуждение,
4) рефлекс.
А4. Рецепторы – это чувствительные образования, которые:
1) передают импульсы в центральную нервную систему,
2) передают нервные импульсы со вставочных нейронов на исполнительные,
3) воспринимают раздражения и преобразуют энергию раздражителей в процесс нервного возбуждения,
4) воспринимают нервные импульсы от чувствительных нейронов.
А5. Наиболее чувствительны к недостатку кислорода клетки:
1) спинного мозга,
2) головного мозга,
3) печени и почек,
4) желудка и кишечника.
А6. Пучки длинных отростков нейронов, покрытые соединительнотканной оболочкой и расположенные вне центральной нервной системы, образуют:
1) нервы,
2) мозжечок,
3) спинной мозг,
4) кору больших полушарий.
А7. Произвольные движения человека обеспечивают:
1) мозжечок и промежуточный мозг,
2) средний и спинной мозг,
3) продолговатый мозг и мост,
4) большие полушария переднего мозга.
А8. Регуляцию и согласование физиологических процессов, протекающих во внутренних органах, обеспечивает:
1) промежуточный мозг,
2) средний мозг,
3) спинной мозг,
4) мозжечок.
А9. Соматическая нервная система, в отличие от вегетативной, управляет работой:
1) скелетных мышц,
2) сердца и сосудов,
3) кишечника,
4) почек.
А10. Нервные импульсы передаются в мозг по нейронам:
1) двигательным,
2) вставочным,
3) чувствительным,
4) исполнительным.
А11. Центры глотательных, дыхательных, сердечно-сосудистых и других жизненно важных рефлексов располагаются в:
1) мозжечке,
2) среднем мозге,
3) продолговатом мозге,
4) промежуточном мозге.
А12. Вегетативная нервная система участвует в:
1) осуществлении произвольных движений,
2) восприятии зрительных, слуховых и вкусовых раздражений,
3) регуляции обмена веществ и работы внутренних органов,
4) формировании звуков речи.
А13. Нервным импульсом называют:
1) электрическую волну, бегущую по нервному волокну,
2) передачу информации с одного нейрона на следующий,
3) передачу информации от клетки к клетке,
4) процесс, обеспечивающий торможение клетки-адресата.
А14. По чувствительному нейрону возбуждение направляется:
1) в центральную нервную систему,
2) к исполнительному органу,
3) к рецепторам,
4) к мышцам.
А15. Нервные импульсы передаются от органов чувств в мозг по:
1) двигательным нейронам,
2) вставочным нейронам,
3) чувствительным нейронам,
4) коротким отросткам двигательных нейронов.
А16. Внешние раздражители преобразуются в нервные импульсы в:
1) нервных волокнах,
2) телах нейронов центральной нервной системы,
3) рецепторах,
4) телах вставочных нейронов.
А17. У человека за расширение зрачка отвечает:
1) симпатический отдел нервной системы,
2) парасимпатический отдел нервной системы,
4) центральная нервная система.
А18. Короткий отросток нервной клетки называется:
1) аксон,
2) нейрон,
3) дендрит,
4) синапс.
А19. Длинный отросток нервной клетки называется:
1) аксон,
2) нейрон,
3) дендрит,
4) синапс.
А20. Место контактов двух нервных клеток друг с другом называется:
1) аксон,
2) нейрон,
3) дендрит,
4) синапс.
А21. Нервы – это:
1) нейронная цепь,
2) скопление тел нейронов,
3) пучки аксонов, выходящие за пределы мозга,
4) рецепторы.
А22. Полушария головного мозга соединяются друг с другом:
1) мостом,
2) мозолистым телом,
3) средним мозгом,
4) промежуточным мозгом.
А23. Влияние парасимпатической нервной системы на сердечную деятельность выражается в:
1) замедлении сердцебиения,
2) учащении сердцебиения,
3) остановке сердца,
4) аритмии.
А24. Нервная система – это:
1) орган,
2) ткань,
3) система органов,
4) органоид.
А25. Нервная система человека, в отличие от эндокринной:
1) реагирует на внешние, а не на внутренние воздействия,
2) полностью подчинена сознанию,
3) действует быстрее,
4) не работает во время сна.
А26. Рефлексы, которые не могут быть усилены или заторможены по воле человека, осуществляются через нервную систему:
1) центральную,
2) вегетативную,
3) соматическую,
4) периферическую.
А27. Аксоны – отростки нервных клеток, которые выходят за пределы центральной нервной системы, собираются в пучки и образуют:
1) подкорковые ядра,
2) нервные узлы,
3) кору мозжечка,
4) нервы.
А28. Нейрон – это:
1) многоядерная клетка с отростками,
2) одноядерная клетка с отростками,
3) безъядерная клетка с отростками,
4) многоядерная клетка с ресничками.
А29. В приспособительных реакциях организма на изменения условий среды ведущую роль играет:
1) головной мозг,
2) вегетативная нервная система,
3) соматическая нервная система,
4) органы чувств.
А30. Нервные клетки отличаются от остальных наличием:
1) ядра с хромосомами,
2) отростков разной длины,
3) многоядерностью,
4) сократимостью.
А31. Передача возбуждения по нерву или мышце объясняется:
1) разностью концентраций ионов натрия и калия внутри и вне клетки,
2) разрывом водородных связей между молекулами воды,
3) изменением концентрации водородных ионов,
4) теплопроводностью воды.
А32. Рефлекс, нервный центр которого лежит за пределами продолговатого мозга:
1) кашель,
2) глотание,
3) слюноотделение,
4) коленный.
А33. Промежуточный мозг регулирует:
1) обмен веществ,
2) потребление пищи и воды,
3) поддержание постоянной температуры тела,
4) верны все ответы.
А34. В продолговатом мозге расположен центр рефлекса:
1) чихания,
2) мочеиспускания,
3) дефекации,
4) коленного.
А35. Центры кашля и чихания находятся в:
1) спинном мозге,
2) продолговатом мозге,
3) среднем мозге,
4) переднем мозге.
А36. Парасимпатическая нервная система снижает:
1) частоту сердечных сокращений,
2) силу сердечных сокращений,
3) уровень глюкозы в плазме,
4) все перечисленные параметры.
Задания с выбором нескольких правильных ответов.
В1. Белое вещество переднего отдела головного мозга:
А) образует его кору,
Б) расположено под корой,
В) состоит из нервных волокон,
Г) образует подкорковые ядра,
Д) соединяет кору головного мозга с другими отделами головного мозга и со спинным мозгом,
Е) выполняет функцию высшего анализатора сигналов от всех рецепторов тела.
В2. Деятельность каких органов регулирует вегетативная нервная система человека?
А) мышц верхних и нижних конечностей,
Б) сердца и кровеносных сосудов,
В) органов пищеварения,
Г) мимических мышц,
Д) почек и мочевого пузыря,
Е) диафрагмы и межрёберных мышц.
В3. К периферической нервной системе относят:
А) мост,
Б) мозжечок,
В) нервные узлы,
Г) спинной мозг,
Д) чувствительные нервы,
Е) двигательные нервы.
В4. В мозжечке лежат центры регуляции:
А) мышечного тонуса,
Б) сосудистого тонуса,
В) позы и равновесия тела,
Г) координации движений,
Д) эмоций,
Е) вдоха и выдоха.
Задания на установление соответствия.
В5. Установите соответствие между отдельной функцией нейрона и типом нейрона, который эту функцию выполняет.
В6. Установите соответствие между отделами нервной системы и их функциями.
| ВЫПОЛНЯЕМЫЕ ФУНКЦИИ | ОТДЕЛ НЕРВНОЙ СИСТЕМЫ |
| 1) сужает сосуды, | А) симпатическая, |
| 2) урежает ритм работы сердца, | Б) парасимпатическая. |
| 3) сужает бронхи, | |
| 4) расширяет зрачок. |
В7. Установите соответствие между строением и функциями нейрона и его отростками.
В8. Установите соответствие между свойствами нервной системы и её типами, которые этими свойствами обладают.
В9. Установите соответствие между примерами нервной деятельности человека и функциями спинного мозга.
В10. Установите соответствие между особенностью строения и функцией головного мозга и его отделом.
Задание на установление правильной последовательности.
В11. Установите правильную последовательность расположения отделов ствола головного мозга, по направлению от спинного мозга.
А) промежуточный мозг,
Б) продолговатый мозг,
В) средний мозг,
Г) мост.
Задания со свободным ответом.
С1. Найдите ошибки в приведённом тексте. Укажите номера предложений, которых сделаны ошибки, объясните их.
1. Кора больших полушарий образована серым веществом.
2. Серое вещество состоит из отростков нейронов.
3. Каждое полушарие разделяется на лобную, теменную, височную и затылочную доли.
4. Зрительная зона находится в лобной доле.
5. Слуховая зона находится в теменной доле.
С2. Найдите ошибки в приведённом тексте. Укажите номера предложений, в которых допущены ошибки, объясните их.
1. Нервная система делится на центральную и соматическую.
2. Соматическая нервная система делится на периферическую и вегетативную.
3. Центральный отдел соматической нервной системы состоит из спинного и головного мозга.
4. Вегетативная нервная система координирует деятельность скелетной мускулатуры и обеспечивает чувствительность.
Ответы к заданиям части А.
| № | ||||||||||||
| ответ | ||||||||||||
| № | ||||||||||||
| ответ | ||||||||||||
| № | ||||||||||||
| ответ |
Ответы к заданиям части В с выбором нескольких правильных ответов.
| № задания | ||||||
| ответ | ||||||
| № задания | ||||||
| ответ |
Похожие статьи
-
Людмила Петрушевская - Странствия по поводу смерти (сборник)
В этой книге собраны истории, так или иначе связанные с нарушениями закона: иногда человек может просто ошибиться, а иногда – посчитать закон несправедливым. Заглавная повесть сборника «Странствия по поводу смерти» – детектив с элементами...
-
Пирожные Milky Way Ингредиенты для десерта
Милки Вэй – очень вкусный и нежный батончик с нугой, карамелью и шоколадом. Название конфеты весьма оригинальное, в переводе означает «Млечный путь». Попробовав его однажды, навсегда влюбляешься в воздушный батончик, который принес...
-
Как оплатить коммунальные услуги через интернет без комиссии
Оплатить услуги жилищно-коммунального хозяйства без комиссий удастся несколькими способами. Дорогие читатели! Статья рассказывает о типовых способах решения юридических вопросов, но каждый случай индивидуален. Если вы хотите узнать, как...
-
Когда я на почте служил ямщиком Когда я на почте служил ямщиком
Когда я на почте служил ямщиком, Был молод, имел я силенку, И крепко же, братцы, в селенье одном Любил я в ту пору девчонку. Сначала не чуял я в девке беду, Потом задурил не на шутку: Куда ни поеду, куда ни пойду, Все к милой сверну на...
-
Скатов А. Кольцов. «Лес. VIVOS VOCO: Н.Н. Скатов, "Драма одного издания" Начало всех начал
Некрасов. Скатов Н.Н. М.: Молодая гвардия , 1994. - 412 с. (Серия "Жизнь замечательных людей") Николай Алексеевич Некрасов 10.12.1821 - 08.01.1878 Книга известного литературоведа Николая Скатова посвящена биографии Н.А.Некрасова,...
-
Кузнецов Виктор Васильевич
При всей известности его острых и прочных ножей в России и за рубежом, часто можно услышать вопросы: когда и где появился на свет Виктор Кузнецов? Биография кузнеца проста и замысловата одновременно. Виктор Васильевич Кузнецов родился в...